
Ringkasan
Pemuliaan tanaman pada dasarnya bergantung pada persilangan meiotik (CO) untuk menata ulang materi genetik dan mengintegrasikan alel yang menguntungkan ke dalam kultivar elit. Frekuensi rekombinasi sangat penting dalam proses ini. Laju rekombinasi yang lebih tinggi meningkatkan kemungkinan pemutusan hambatan hubungan dan menghasilkan kombinasi alel baru. Di sini, dengan menggunakan padi ( Oryza sativa ) sebagai spesies tanaman model, kami mengungkap bahwa RECQ4, penekan yang dilestarikan dari pembentukan CO meiotik, sangat diperlukan untuk menjaga integritas metabolisme intermediet rekombinasi meiotik. Kami menunjukkan bahwa RECQ4 membatasi CO pada padi dengan secara khusus menekan jalur CO yang tidak mengganggu. Redundansi genetik dengan FANCM menggarisbawahi fungsi kooperatif mereka dalam memastikan pembentukan CO kanonik, yang penting untuk segregasi homolog yang akurat dan stabilitas genom. Lebih jauh, mutan ganda recq4 dmc1 menunjukkan fragmentasi kromosom yang persisten, yang melibatkan RECQ4 dalam menyelesaikan intermediet rekombinasi melalui perbaikan kromatid saudara. Temuan kami mendefinisikan ulang peran RECQ4 dalam meiosis tanaman, menjembatani aktivitas penekanan CO dengan fungsi pengawasan genom yang lebih luas.
Perkenalan
Rekombinasi meiosis merupakan proses mendasar dalam reproduksi seksual. Pertama, pembentukan crossover (CO) sangat penting untuk pemisahan kromosom yang akurat selama pembentukan gamet. Kedua, pertukaran materi genetik timbal balik antara kromosom homolog memfasilitasi penataan ulang informasi genetik parental, memastikan bahwa materi rekombinasi diwariskan ke generasi berikutnya. Hal ini menjadikan pembentukan CO meiosis sebagai target utama dalam pemuliaan tanaman. Meiosis dimulai dengan pengenalan sejumlah kerusakan untai ganda DNA (DSB), yang sebagian besar diperbaiki melalui peristiwa non-CO dan hanya sedikit di antaranya yang mengarah pada pembentukan kromosom rekombinan.
Rekombinasi meiotik dimulai oleh DSB terprogram, yang dikatalisis oleh protein SPO11 dan MTOPVIB (Keeney et al ., 1997 ; Robert et al ., 2016 ; Xue et al ., 2016 ). DSB ini kemudian diproses melalui reseksi untai, membentuk ekor DNA untai tunggal berujung 3′ (ssDNA). Ekor ssDNA ini segera diikat oleh protein RPA, yang kemudian digantikan oleh DMC1 atau RAD51 untuk menyerang sekuens homolog.
Baik rekombinase RAD51 maupun DMC1 mengkatalisis perbaikan kerusakan meiosis, memastikan terjadinya CO antara kromosom homolog (inter-homolog, IH) dan bukan antara kromatid saudara yang berdekatan (inter-sister, IS). Penelitian terkini, yang terutama difokuskan pada meiosis budding yeast, menunjukkan bahwa perbaikan kerusakan meiosis terjadi dalam dua fase temporal yang berbeda: fase awal yang memungkinkan DMC1 (Fase 1), diikuti oleh fase yang memungkinkan RAD51 (Fase 2) (Ziesel et al ., 2022 ). Selama fase permisif DMC1, DMC1 terutama memperbaiki DSB dan memfasilitasi rekombinasi antar-homolog, sementara RAD51 tetap tidak aktif secara katalitik (Tsubouchi & Roeder, 2006 ; Busygina et al ., 2008 ; Niu et al ., 2009 ; Lao et al ., 2013 ; Callender et al ., 2016 ). Dalam fase permisif RAD51, jalur yang dimediasi RAD51 menjadi aktif, memperbaiki DSB yang tersisa, terutama menggunakan kromatid saudara (Crismani et al ., 2013 ; Enguita-Marruedo et al ., 2019 ; Toraason et al ., 2021 ; Emmenecker et al ., 2024 ). Namun, pada tumbuhan, RAD51 dapat memperbaiki kerusakan tanpa adanya DMC1, meskipun ia melakukannya dengan menggunakan kromatid saudara, sebagaimana dibuktikan oleh hampir tidak adanya CO antar-homolog (Couteau et al ., 1999 ; Wang et al ., 2016 ).
Dua jenis CO yang berbeda diketahui. Pembentukan CO Kelas I bergantung pada sekumpulan protein spesifik meiosis yang dikenal sebagai ZMM (ZIP1, ZIP2, ZIP3, ZIP4, MER3, MSH4, PTD, HEIP1, dan MSH5) (Li et al ., 2018 ; Li et al ., 2023 ; Luo et al ., 2013 ; Shen et al ., 2012 ; Wang et al ., 2009 ; Wang et al ., 2010 ; Wang et al ., 2012 ; Zhang et al ., 2014 ). CO ini menunjukkan interferensi, yang berarti mereka berjarak lebar di sepanjang kromosom. Pada organisme seperti budding yeast dan Arabidopsis , CO Kelas I mencakup 80%–90% dari total CO (Lambing et al ., 2017 ; Osman et al ., 2011 ). CO yang tersisa, yang diklasifikasikan sebagai Kelas II, sebagian bergantung pada endonuklease MUS81 (Higgins et al ., 2008 ; Mu et al ., 2023 ; Osman et al ., 2011 ).
Bahasa Indonesia: Dalam Arabidopsis , penelitian ekstensif oleh Mercier dan koleganya telah mengidentifikasi beberapa faktor yang menghambat CO [anti-crossover (anti-CO)] selama meiosis, khususnya yang membatasi pembentukan CO Kelas II (Crismani et al ., 2012 ; Fernandes et al ., 2018a , 2018b ; Girard et al ., 2014 ; Séguéla-Arnaud et al ., 2015 , 2017 ; Serra et al ., 2018 ). Faktor-faktor anti-CO ini dikelompokkan ke dalam tiga kategori utama: helicase FANCM dan kofaktornya (MHF1 dan MHF2), kompleks BLM-TOP3α–RMI dan kompleks FIGNL1-FLIP. Faktor-faktor ini memiliki efek aditif pada pembentukan CO tetapi memiliki sedikit atau tidak ada dampak pada fertilitas. Penelitian selanjutnya pada berbagai tanaman (Arrieta et al ., 2021 ; Blary et al ., 2018 ; Desjardins et al ., 2022 ; Li et al ., 2021 , 2023 ; Mieulet et al ., 2018 ; Osman et al ., 2024 ), termasuk tanaman seperti gandum, padi, tomat, dan kacang polong, telah mengonfirmasi sifat lestari dari fungsi gen ini, sementara juga mengungkap beberapa perbedaan khusus spesies, yang menunjukkan potensi divergensi dalam perannya. Selain itu, hilangnya fungsi RPA1a padi mengakibatkan pelepasan represi yang signifikan pada CO Kelas II, meskipun hal ini disertai dengan kemandulan (Miao et al ., 2021 ).
Peningkatan laju CO dapat meningkatkan kombinasi sifat-sifat menguntungkan dari berbagai garis induk, sehingga meningkatkan efisiensi pemuliaan. Beras, tanaman pokok yang menjadi sumber makanan bagi lebih dari setengah populasi dunia, sangat penting untuk mengatasi tantangan yang terkait dengan pertumbuhan populasi dan tekanan lingkungan, seperti perubahan iklim. Mengembangkan varietas padi yang lebih baik sangat penting untuk memastikan ketahanan pangan. Meskipun peningkatan laju CO telah didokumentasikan pada mutan RECQ4 padi (Mieulet et al ., 2018 ), mekanisme molekuler yang mendasari fenotipe ini masih kurang dipahami. Yang terpenting, interaksi fungsional antara RECQ4 dan helicase FANCM, khususnya regulasi terkoordinasi mereka terhadap pembentukan CO Kelas II, belum dikarakterisasi pada spesies ini.
Hasil
RECQ4 tidak diperlukan untuk kesuburan padi
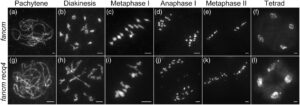
Untuk menyelidiki lebih lanjut fungsi meiosis RECQ4, kami memanfaatkan sistem clustered regular interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) untuk menghasilkan mutasi spesifik pada gen ini. Tiga galur transgenik independen diperoleh pada varietas padi Zhongxian 3037 (Gambar 1 ). Ketiga mutan menunjukkan pertumbuhan vegetatif dan viabilitas serbuk sari yang normal dibandingkan dengan tipe liar (Gambar 1a,b ). Di antara ini, kami memilih recq4-1 untuk analisis lebih lanjut (Gambar 1c,d ). Pada tipe liar, kromosom mengalami kondensasi untuk membentuk benang panjang dan tipis selama leptoten. Melalui zigoten dan pakiten, kromosom homolog berpasangan dan bersinaps. Dari diploten ke diakinesis, setelah kompleks sinaptonemal dibongkar, CO antara kromosom homolog matang menjadi kiasmata yang terlihat, kromosom semakin terkondensasi dan 12 bivalen diamati dengan jelas. Bivalen berjajar secara teratur pada lempeng ekuator selama metafase I, diikuti oleh pemisahan dan migrasi kromosom homolog dalam arah yang berlawanan selama anafase I. Pada pembelahan meiosis kedua, kromatid saudara dari setiap kromosom terpisah, menghasilkan empat sel anak, masing-masing berisi 12 kromosom (Gambar 1e ). Meiosis jantan pada mutan recq4 tidak menunjukkan cacat yang terdeteksi, konsisten dengan fertilitas tipe liar.
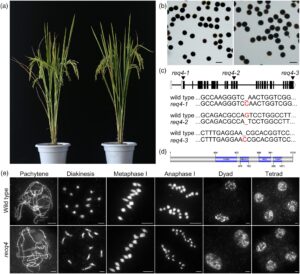
Mutasi RECQ4 meningkatkan pembentukan bivalen pada mutan zmm padi
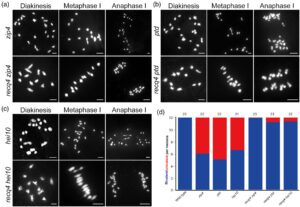
Aktivitas anti-CO meiosis RECQ4 awalnya dikarakterisasi melalui penyaringan genetik, karena mutasinya memulihkan pembentukan bivalen pada mutan ZMM di Arabidopsis (Arrieta et al ., 2021 ). Kami selanjutnya menyelidiki hubungan antara ZMM dan RECQ4 pada monokotil, khususnya pada padi. Mutan ganda recq4 zmm dihasilkan, dan kami mengukur jumlah bivalen di samping mutan hei10 , ptd , dan zip4 . Ini dibandingkan dengan jumlah bivalen pada mutan recq4 hei10 , recq4 ptd, dan recq4 zip4 dengan mempelajari konfigurasi kromosom pada metafase I. Mutan ganda recq4 zmm menunjukkan frekuensi bivalen yang meningkat secara signifikan dibandingkan dengan mutan ZMM saja dengan genotipe yang sesuai. Pada mutan ganda recq4 zmm , 12 bivalen terdeteksi secara mudah, sedangkan yang tidak teramati pada mutan ZMM , hal ini menunjukkan bahwa RECQ4 membatasi pembentukan CO meiosis (Gambar 2 ).
Pembentukan DSB, pengikatan ssDNA-DMC1 dan perakitan SC tampak normal pada recq4
Untuk menentukan apakah RECQ4 memiliki dampak pada jumlah DSB, kami melakukan studi imunolokalisasi dengan anti-γH2AX pada meiosit recq4 . Jumlah rata-rata fokus γH2AX adalah 217 ± 30 ( n = 26) pada recq4 dan 223 ± 36 ( n = 17) pada tipe liar pada tahap yang sama dan tidak berbeda secara signifikan ( uji- t tidak berpasangan , P = 0,60), yang menunjukkan bahwa pembentukan DSB terjadi secara normal pada recq4 (Gambar 3a ). Untuk mengonfirmasi hal ini lebih lanjut, kami menghasilkan mutan ganda recq4 spo11-2 (Gambar S1 ), yang tampak mirip dengan fenotipe mutan tunggal spo11-2 dengan 24 univalen yang tersebar secara acak selama pembelahan meiosis pertama (Gambar 3b ). Oleh karena itu, RECQ4 berfungsi sebagai respons terhadap DSB meiosis.
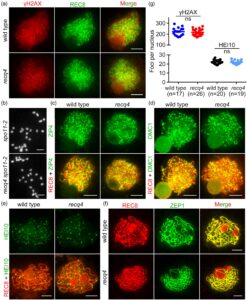
Lebih jauh, kami melakukan imunopewarnaan menggunakan antibodi terhadap DMC1, rekombinase spesifik meiosis, yang memediasi pertukaran untai homolog (Gambar 3d ; Gambar S2 ). Jumlah fokus DMC1 tidak berbeda secara signifikan ( uji t tidak berpasangan , P = 0,05) pada mutan recq4 (239 ± 9, n = 24) dari yang ada pada tipe liar (264 ± 9, n = 26) pada zigoten, yang menunjukkan bahwa peningkatan CO tidak terkait dengan peningkatan DSB.
Berdasarkan pewarnaan 4′,6-diamidino-2-phenylindole (DAPI), sinapsis kromosom homolog pada dasarnya normal pada meiosit recq4 (Gambar 1e ). Untuk mengonfirmasi pembentukan SC pada mutan recq4 , kami melakukan imunolokalisasi dengan antibodi terhadap ZEP1, protein filamen transversal SC. Lokalisasi ZEP1 tidak dapat dibedakan dari tipe liar pada pachytene (Gambar 3f ), yang menunjukkan bahwa pembentukan SC telah selesai pada recq4 .
recq4 menunjukkan jumlah fokus HEI10 terang yang normal
Jalur peka-interferensi (CO I), yang bergantung pada protein ZMM (seperti ZIP4), merupakan penyebab sebagian besar CO pada padi. Untuk memverifikasi frekuensi pembentukan CO peka-interferensi pada recq4 , pertama-tama kami menentukan lokalisasi ZIP4 pada tanaman tipe liar dan mutan recq4 . Distribusi ZIP4 pada recq4 mirip dengan yang ada pada tipe liar (Gambar 3c ), yang menunjukkan bahwa CO yang bergantung pada ZMM terbentuk secara normal. Selanjutnya, kami menemukan bahwa jumlah rata-rata fokus terang HEI10 dari recq4 (21 ± 2, n = 19) tidak berbeda secara signifikan ( uji- t tidak berpasangan , P = 0,55) dibandingkan dengan tipe liar (22 ± 2, n = 20). Oleh karena itu, pembentukan CO peka-interferensi berlangsung secara normal pada recq4 selama meiosis padi (Gambar 3e,g ).
MUS81 diperlukan untuk menyelesaikan perantara rekombinasi di recq4
Kurangnya viabilitas mutan ganda recq4 mus81 di Arabidopsis membatasi penelitian lebih lanjut tentang meiosis yang melibatkan genotipe ini dalam laporan sebelumnya (Arrieta et al ., 2021 ). Untungnya, kami memperoleh mutan recq4 mus81 pada padi. Meskipun menunjukkan cacat pertumbuhan yang ditandai dengan tinggi tanaman yang lebih rendah (Gambar S3 ), meiosit dapat diperoleh dari tanaman recq4 mus81 . Perkembangan meiosis dari leptoten ke metafase I tampak normal, sebagaimana dibuktikan oleh pewarnaan DAPI pada kromosom meiosis di PMC (Gambar 4a ). Deteksi fragmentasi kromosom selama anafase I sesuai dengan sterilitas yang diamati, sebuah fenomena yang tidak ada pada mutan tunggal recq4 atau mus81 (Mu et al ., 2023 ). Pengamatan ini menunjukkan bahwa MUS81 menjadi sangat diperlukan untuk perbaikan komprehensif perantara rekombinasi di recq4 .
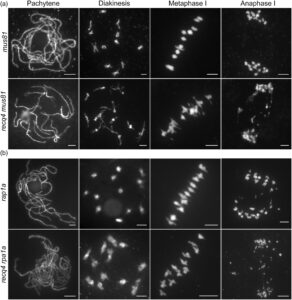
Perilaku kromosom pada mutan ganda rpa1a recq4 merangkum fenotipe mutan tunggal rpa1a
Penelitian kami sebelumnya telah menunjukkan bahwa protein perancah RPA1a memainkan peran penting dalam membatasi pembentukan kiasma yang berlebihan selama meiosis padi (Miao et al ., 2021 ). Fungsi ini sebagian dikaitkan dengan aktivitas abnormal homolog protein sindrom Bloom (BLM), RECQ4, dalam kompleks BLM-TOP3α–RMI. Berdasarkan temuan ini, kami menduga mutasi pada RECQ4, anggota kompleks ini, pada latar belakang rpa1a tidak akan menyebabkan perubahan perilaku kromosom dibandingkan dengan mutan tunggal rpa1a . Untuk menemukan perbedaan morfologi kromosom di antara dua mutan tunggal, recq4 dan rpa1a , dan mutan ganda recq4 rpa1a , kami menyelidiki dengan hati-hati perilaku kromosom mereka selama anafase I. recq4 dekat dengan tipe liar tanpa fragmen kromosom apa pun pada anafase I, sementara rpa1a mirip dengan mutan ganda, semuanya dengan fragmen kromosom pada anafase I (Gambar 4b ; Gambar S1 ). Dengan demikian, kami berhipotesis bahwa RPA1A dan RECQ4 berfungsi untuk membatasi pembentukan CO yang tidak mengganggu, tetapi RPA1A kemungkinan bertindak di hulu RECQ4 dalam jalur pengaturan ini.
Tumpang tindih fungsional antara RECQ4 dan FANCM memastikan segregasi kromosom meiosis yang akurat selama meiosis padi
Frekuensi CO meningkat sembilan kali lipat pada mutan ganda Arabidopsis recq4 fancm (Séguéla-Arnaud et al ., 2015 ), yang mendorong kami untuk mengeksplorasi fungsi meiosis mutan ganda recq4 fancm pada spesies tanaman pangan. Kami menghasilkan mutan ganda pada padi menggunakan dua metode: teknologi CRISPR dan hibridisasi. Mutasi ganda recq4 fancm menunjukkan pertumbuhan vegetatif normal tetapi hampir steril (Gambar S4 ). Untuk menyelidiki penyebab yang mendasari sterilitas pada mutan recq4 fancm , kami menganalisis kromosom meiosis sel induk serbuk sari pada berbagai tahap meiosis baik pada tipe liar maupun mutan recq4 fancm .
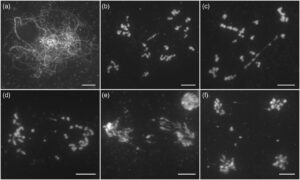
Perilaku kromosom mutan ganda recq4 fancm tidak dapat dibedakan dari tipe liar dari leptotene hingga pakitene. Namun, anomali mulai terlihat setelahnya (Gambar 5 ). Tidak seperti 12 bivalen kondensasi utuh yang diamati pada mutan tipe liar atau fancm (Gambar 5b ), interaksi kromosom abnormal ini menjadi lebih jelas pada diakinesis atau metafase I (Gambar 5h,i ), di mana kromosom muncul sebagai massa yang terjerat, meskipun struktur seperti bivalen masih ada (Gambar 5i ). Selama anafase I, jembatan dan fragmen kromosom yang luas diamati pada pelat (Gambar 5g ). Setelah meiosis II (Gambar 5k ), tetrad dengan distribusi kromosom dan mikronukleus yang tidak sama terbentuk (Gambar 5l ). Cacat serupa diamati selama meiosis pada mutan ganda penyisipan T-DNA recq4 fancm (Gambar S5 ). Data ini menggarisbawahi redundansi genetik antara RECQ4 dan FANCM, yang menjaga pemisahan kromosom meiotik yang akurat.
Kehilangan fungsi pada RECQ4 dan FANCM menunjukkan kapasitas yang lebih besar untuk menyelamatkan pembentukan bivalen pada latar belakang kekurangan ZMM
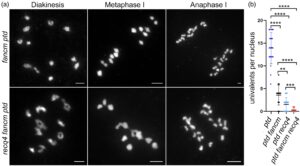
Kami menghasilkan mutan rangkap tiga (TM) recq4 fancm ptd untuk memeriksa apakah pembentukan univalen lebih jauh berkurang dibandingkan dengan mutan ganda recq4 ptd menggunakan penilaian sitologi (Gambar 6 ; Gambar S1 ). Kami mengamati penurunan signifikan ( uji- t tak berpasangan , P < 0,01) dalam pembentukan univalen di TM recq4 fancm ptd (0,29 ± 0,09 per sel, n = 24), dibandingkan dengan mutan recq4 ptd (1,55 ± 0,32 per sel, n = 22) dan mutan ganda fancm ptd (3,58 ± 0,50 per sel, n = 19) (Gambar 6b ). Akibatnya, kami menduga bahwa mutasi kehilangan fungsi di RECQ4 dan FANCM dapat menunjukkan kapasitas lebih besar untuk menyelamatkan pembentukan bivalen di latar belakang defisiensi ZMM.
RECQ4 mungkin terlibat dalam perbaikan DSB meiotik melalui kromatid saudara
Peran RECQ4 dalam konteks defisiensi DMC1 belum dipelajari secara ekstensif. Untuk menyelidiki fungsinya, kami menghasilkan mutan ganda recq4 dmc1 . Analisis pewarnaan DAPI dari mutan ganda tersebut mengungkap sinapsis abnormal dari kromosom homolog, meskipun beberapa daerah dengan penyelarasan yang tepat tetap terlihat (Gambar 7a ). Saat kondensasi berlangsung, sejumlah besar univalen berbentuk tidak teratur diamati pada tahap diakinesis (Gambar 7b ). Univalen ini gagal untuk menyelaraskan dengan benar di sepanjang lempeng ekuator dan tetap tersebar di seluruh nukleus (Gambar 7c ). Selama anafase I, semua kromosom terpisah ke kutub, dengan fragmen kromosom terdeteksi (Gambar 7d ). Fragmen kromosom secara konsisten diamati selama telofase I berikutnya (Gambar 7e ) dan tahap tetrad (Gambar 7f ). Berdasarkan temuan ini, kami dengan hati-hati menyimpulkan bahwa RECQ4 penting untuk perbaikan DSB melalui kromatid saudara.
Diskusi
Diferensiasi fungsi RECQ4 antar spesies
Arabidopsis RECQ4A dan RECQ4B berasal dari duplikasi segmen kromosom baru-baru ini dalam famili Brassicaceae , yang mengarah pada pembentukan dua gen dengan urutan protein yang sangat mirip. Mutasi ganda recq4a recq4b mampu memulihkan pembentukan bivalen dan fertilitas dalam latar belakang zmm . Meskipun terdapat peningkatan jarak genetik sebanyak 6,2 kali lipat, peningkatan rekombinasi tidak mengganggu segregasi kromosom atau fertilitas pada mutan recq4a recq4b (Séguéla-Arnaud et al ., 2015 ). Barli, tomat, dan beras masing-masing hanya memiliki satu gen homolog. Mutasi HvRECQL4 menghasilkan pemulihan fertilitas paling lengkap pada mutan barli semi-fertil mlh3 . Tingkat rekombinasi pada galur HvRECQL4 homozigot hampir dua kali lipat dari tipe liar barli. de Maagd et al . pertama kali ditunjukkan pada tomat bahwa recq4 meningkatkan frekuensi rekombinasi pada hibrida interspesifik (de Maagd et al ., 2020 ). Mutasi biallelik recq4 dibuat pada hibrida F1 Solanum lycopersicum dan S. pimpinellifolium . Dibandingkan dengan hibrida F1 tipe liar, mutan recq4 menunjukkan peningkatan signifikan pada CO: peningkatan 1,53 kali lipat pada bivalen cincin yang diamati secara mikroskopis pada meiosit jantan, dan perluasan 1,8 kali lipat peta genetik berdasarkan analisis penanda SNP pada keturunan F2. Melalui analisis sitologi, penelitian kami secara independen mengonfirmasi bahwa RECQ4 membatasi CO meiosis pada padi. Pada gandum ( Triticum aestivum L.) TM yang tidak memiliki tiga salinan homoeolog TaRECQ4, pengamatan fenotipik mengungkapkan penurunan signifikan pada fertilitas dan viabilitas serbuk sari. Kehadiran sejumlah multivalen pada tanaman TM menunjukkan bahwa TaRECQ4 mungkin juga berperan dalam mengatur rekombinasi homolog (Bazile et al ., 2023 ). Berdasarkan hasil di atas, peran RECQ4 dalam allopolyploid layak untuk dieksplorasi lebih lanjut.
Seberapa tinggikah laju rekombinasi selama meiosis padi?
Arabidopsis , model tumbuhan dikotil dengan genom terkecil, menunjukkan efek terbesar saat menggabungkan mutasi recq4 dan figl1 . Hal ini menghasilkan peningkatan signifikan dalam panjang peta genetik hibrida, dari 389 menjadi 3037 cM, yang sesuai dengan peningkatan frekuensi CO yang belum pernah terjadi sebelumnya sebanyak 7,8 kali lipat (Fernandes et al ., 2018a ). Pengacauan lebih lanjut pada ketiga jalur tersebut tidak menyebabkan peningkatan rekombinasi tambahan, yang menunjukkan bahwa batas atas telah tercapai.
Di sini, kami telah melakukan beberapa uji anti-CO pada padi (Hu et al ., 2017 ; Li et al ., 2023 ; Yang et al ., 2022 ). Mengingat redundansi fungsional antara RECQ4 dan FANCM, temuan kami mendukung kesimpulan bahwa manipulasi RECQ4 dapat berfungsi sebagai alat universal untuk meningkatkan rekombinasi pada tanaman (Mieulet et al ., 2018 ), dengan pengecualian gandum allopolyploid (Bazile et al ., 2023 ). Namun, penting untuk dicatat bahwa data yang diperoleh dari spesies model mungkin tidak selalu berlaku langsung pada tanaman pangan.
Kami berspekulasi bahwa perbedaan signifikan antara kesimpulan penelitian ini dan kesimpulan pada Arabidopsis mungkin berasal dari ukuran genom padi yang jauh lebih besar, yang tampaknya terkait dengan frekuensi DSB DNA yang lebih tinggi per sel. Pada mutan ganda Arabidopsis recq4 fancm , mesin perbaikan DNA dapat secara efektif mengelola jumlah perantara rekombinasi abnormal yang relatif rendah, sehingga menghasilkan lebih banyak CO dan lebih sedikit molekul DNA yang tidak diperbaiki. Namun, pada padi, peningkatan jumlah perantara rekombinasi abnormal mengharuskan setidaknya satu dari FANCM atau RECQ4 tetap berfungsi.
Jangan lupakan saudara perempuan Anda: Keterlibatan dalam perbaikan DSB selama meiosis padi
Salah satu ciri khas meiosis adalah pembentukan CO antara kromosom homolog. Model perbaikan kerusakan untai ganda meiosis (DSBR) terutama menyoroti perbaikan IH (An et al ., 2003 ), namun perbaikan kromatid saudara (IS), yang memastikan kesetiaan genomik melalui rekombinasi homolog antara kromatid saudara, belum diselidiki secara memadai (Humphryes dan Hochwagen, 2014 ).
Pilihan pola meiosis (IH vs. IS) bergantung pada regulasi rekombinase RAD51 dan DMC1 yang tepat untuk memastikan keragaman dan kesetiaan genetik. Pada mutan ganda recq4 dmc1 , fragmentasi kromosom yang luas diamati di semua sel dari anafase I hingga sisa meiosis, suatu fenotipe yang tidak ada pada mutan tunggal dmc1 . Perbedaan ini menyiratkan bahwa RECQ4 juga dapat berfungsi dalam DSBR meiosis melalui jalur yang bergantung pada kromatid saudara.
Bahan dan metode
Bahan tanaman
Mutan meiosis yang digunakan dalam penelitian ini, zip4 (Shen et al ., 2012 ), ptd (Li et al ., 2023 ), hei10 (Wang et al ., 2012 ), mus81 (Mu et al ., 2023 ), spo11-2 (Miao et al ., 2021 ), dmc1 (Wang et al ., 2016 ), dan rpa1a (Miao et al ., 2021 ), sebelumnya diisolasi di laboratorium kami. Mutan ganda yang menggabungkan ini dengan recq4 dihasilkan melalui persilangan. Mutan ganda fancm , recq4 fancm (Gambar S1 ) dihasilkan dalam varietas padi Zhongxian 3037 melalui mutagenesis yang dimediasi CRISPR/Cas9 (Wang et al ., 2015 ). Mutan T-DNA dari FANCM (5A-00166) dan RECQ4 (3A-01037), juga digunakan untuk mengkonstruksi mutan ganda, bersumber dari POSTECH Rice Insertion Database (An et al ., 2003 ). TM recq4 fancm ptd dihasilkan dengan memasukkan mutasi ke dalam gen RECQ4 dan FANCM menggunakan penyuntingan gen CRISPR/Cas9 pada keturunan tanaman heterozigot PTD yang melakukan penyerbukan sendiri. Tanaman heterozigot ini berasal dari varietas padi Yandao 8. Genotipe TM ( recq4 fancm ptd ) dikonfirmasi melalui sekuensing PCR. Semua tanaman ditanam di lahan percobaan di Beijing atau Hainan dalam kondisi pertumbuhan normal.
Persiapan kromosom meiotik
Larutan Carnoy (etanol dan asam asetat glasial dengan perbandingan 3:1) digunakan untuk memfiksasi malai muda (6 ± 1 cm) dari tanaman tipe liar dan mutan. Mikrosporosit dari malai muda dipilih, dan kepala sari diremas dalam larutan asetokarmin di bawah penutup kaca. Setelah dibekukan dalam nitrogen cair, penutup kaca segera dilepas (dalam waktu 1 menit), dan slide yang berisi kromosom didehidrasi melalui serangkaian pencucian alkohol. Seri alkohol terdiri dari etanol absolut dan air deionisasi steril dalam konsentrasi berikut: etanol 70%, 90% dan 100% (Cheng, 2013 ). Kromosom pada slide diwarnai dengan DAPI dalam larutan antifade (Vector Laboratories, Burlingame, CA), dan kromosom diamati dan divisualisasikan menggunakan mikroskop fluoresensi ZEISS A2 yang dilengkapi dengan kamera mikro-CCD.
Imunofluoresensi
Larutan paraformaldehida 4% (b/v) digunakan untuk memfiksasi malai muda dari tanaman tipe liar dan mutan. Setelah fiksasi pada suhu ruangan (20–25 °C) selama 30 menit, kepala sari diremas dengan jarum bedah dalam larutan PBS dan ditutup dengan penutup kaca. Kaca objek yang berisi kromosom kemudian dibekukan dalam nitrogen cair. Setelah penutup kaca dilepaskan, kaca objek didehidrasi melalui serangkaian pencucian etanol (70%, 90% dan 100%) (Cheng, 2013 ). Kaca objek kemudian diinkubasi dalam ruang lembap dengan kombinasi antibodi yang berbeda, masing-masing diencerkan 1:500 dalam buffer 1× TNB (0,1 M Tris–HCl, pH 7,5, 0,15 M NaCl dan 0,5% reagen pemblokiran) pada suhu 37 °C selama 4 jam. Setelah tiga putaran pencucian PBS, antibodi antikelinci terkonjugasi Texas-Red dan antibodi antitikus terkonjugasi fluorescein isothiocyanate (FITC) (1:1000) diaplikasikan pada slide. Kromosom diwarnai dengan DAPI dalam larutan antifade (Vector Laboratories). Semua gambar diambil menggunakan mikroskop Carl Zeiss Axio Imager A2 yang dilengkapi dengan kamera mikro-CCD.
Penargetan CRISPR-Cas9 pada RECQ4 dan FANCM
Urutan targetnya adalah GACTTCGACTGGGAGGCGG dan TGATAAAGCCAAGGGTCAACTGG. Vektor biner CRISPR-Cas9 pC1300-cas9 digunakan untuk transformasi genetik.
Analisis statistik
Signifikansi statistik ditentukan oleh uji- t dua sisi yang tidak berpasangan , kecuali dinyatakan lain. GraphPad Prism 6 ( http://www.graphpad.com/ ) digunakan untuk membuat grafik.



Tinggalkan Balasan