Abstrak
Rami industri ( Cannabis sativa L.) adalah tanaman yang muncul kembali di Amerika Serikat dengan tantangan agronomi unik yang memerlukan studi dan panduan khusus lokasi. Alat pertanian digital, seperti model pertumbuhan tanaman, dapat memfasilitasi proses ini dengan memungkinkan pemahaman yang lebih baik tentang sistem pertanian. Model pertumbuhan tanaman memprediksi pertumbuhan dan perkembangan tanaman dari waktu ke waktu dengan menggunakan cuaca, tanah, manajemen, dan parameter fisiologis sebagai masukan. Tujuan dari penelitian ini adalah untuk mengembangkan model rami baru dalam modul Cropping System Model (CSM)-CROPGRO dalam Decision Support System for Agrotechnology Transfer (DSSAT). Data eksperimen yang mencakup dua kultivar, keduanya tumbuh selama dua musim dan dua lokasi di Florida, digunakan untuk kalibrasi dan evaluasi model. Adaptasi model dilakukan dalam (1) komposisi jaringan dan partisi asimilasi, (2) suhu kardinal untuk proses pertumbuhan dan perkembangan yang berbeda, dan (3) fotosintesis dan penuaan daun. Hasilnya menunjukkan simulasi yang baik dari biomassa di atas tanah ( d = 0,91, root mean square error [RMSE] = 482 kg ha −1 ), berat batang ( d = 0,83, RMSE = 430 kg ha −1 ), dan waktu berbunga (+4 hingga −5 hari), yang menangkap perbedaan di antara kultivar dan tanggal tanam. Analisis musiman menggunakan model yang diadaptasi menunjukkan dampak dari tanggal tanam variabel pada fenologi rami, biomassa, dan produksi biji-bijian. Pekerjaan di masa depan harus mencakup pengamatan yang lebih rinci dan simulasi mekanistik penjarangan sendiri dan evaluasi dengan data yang mewakili lingkungan produksi dan kultivar yang berbeda. Model CROPGRO-Hemp akan memberikan dasar bagi petani, peneliti, dan pemangku kepentingan lainnya untuk menganalisis secara sistematis sistem produksi rami di Florida dan internasional.
Ringkasan Bahasa Sederhana
Rami industri adalah tanaman yang ditanam untuk serat dan benih, dengan minat baru di Amerika Serikat karena banyaknya kegunaan dan manfaat berkelanjutannya. Studi ini mengembangkan dan menguji model komputer baru untuk memprediksi bagaimana rami tumbuh dan menghasilkan serat dan benih di Florida. Model ini membantu petani dan peneliti memahami bagaimana tanggal penanaman, lokasi, dan jenis tanaman yang berbeda memengaruhi pertumbuhan rami. Model tersebut secara akurat memprediksi pertumbuhan rami, waktu berbunga, dan hasil panen menggunakan data lapangan. Model tersebut menunjukkan bahwa menanam rami sekitar tanggal 1 Juni di Florida utara memberikan hasil panen serat tertinggi, karena respons tanaman terhadap panjang hari. Model tersebut juga menemukan bahwa siang hari yang lebih panjang atau jenis rami khusus dapat sangat meningkatkan produktivitas. Model rami baru adalah alat yang berharga untuk merencanakan dan meningkatkan pertanian rami di Florida dan sekitarnya. Memilih tanggal penanaman dan jenis rami yang tepat sangat penting untuk produksi terbaik. Model tersebut akan membantu memandu penelitian dan keputusan pertanian di masa mendatang.
Singkatan
Bahasa Indonesia: CSM
Model Sistem Tanam
Sistem Penyiaran Digital (DSS)
sistem pendukung keputusan
Bahasa Indonesia: DSSAT
Sistem Pendukung Keputusan untuk Transfer Agroteknologi
LAI
indeks luas daun
MSE
kesalahan kuadrat rata-rata
PAR
radiasi aktif fotosintesis
PD
hari fototermal
RMSE
kesalahan akar kuadrat rata-rata
MENYESALI
efisiensi penggunaan radiasi
1. PENDAHULUAN
Rami industri ( Cannabis sativa L.) adalah tanaman dioecious tahunan dengan berbagai kegunaan untuk serat, biji, minyak, dan keperluan pengobatan. Tanaman ini termasuk dalam famili Cannabaceae, bersama dengan hop ( Humulus lupulus L.) dan hackberry ( Celtis spp.). Rami memiliki sejarah panjang dalam budidaya dan penggunaan, yang dimulai sejak 8000 tahun yang lalu di Tiongkok, India, dan Timur Tengah (Small, 2015 ). Rami dianggap sebagai alternatif yang lebih berkelanjutan dan hemat air untuk serat kapas ( Gossypium L.) (Schumacher et al., 2020 ; Wise et al., 2023 ) dan telah digunakan dalam berbagai aplikasi industri mulai dari manufaktur hingga konstruksi (Ranalli & Venturi, 2004 ; Rehman et al., 2021 ; Van der Werf et al., 1996 ). Mengingat perubahan kebijakan terkini dan legalisasi tanaman ini, rami kembali menarik minat, terutama di Amerika Utara. Selanjutnya, beberapa program penelitian regional dibentuk (Cherney & Small, 2016 ; Mylavarapu et al., 2020 ) untuk mempelajari produksi dan ekonomi rami, tetapi juga aspek sosial-ekologisnya seperti risiko invasi dan persepsi konsumen (Canavan et al., 2022 ; Rampold et al., 2021 ). Luas lahan terbuka untuk rami serat, biji, dan gandum pada tahun 2023 di Amerika Serikat mencapai 7056 ha dan memiliki nilai $16,82 juta, peningkatan signifikan dibandingkan tahun 2022 (USDA NASS, 2024 ). Meskipun kegunaannya dan pasarnya sangat luas (Expert Market Research, 2024 ; G. Kaur & Kander, 2023 ), banyak penelitian menyoroti keterbatasan pengetahuan dan kebutuhan penelitian saat ini mengenai produksi dan ekonomi rami (Adesina et al., 2020 ; Mark et al., 2020 ).
Di sisi agronomi, memahami variabilitas dan interaksi faktor genetik, lingkungan, dan manajemen merupakan langkah penting saat memperkenalkan tanaman baru untuk mengidentifikasi atau mengembangkan kultivar yang beradaptasi secara lokal dan pendekatan manajemen (Hatfield & Walthall, 2015 ). Mengingat kompleksitas interaksi ini, ilmu data dan pemodelan pertumbuhan tanaman berbasis proses adalah pendekatan untuk memajukan pemahaman kita tentang tanaman rami dan sistem pertanian di sekitarnya (Jones et al., 2017 ; Tsuji et al., 1998 ). Model pertumbuhan tanaman mensimulasikan pertumbuhan dan perkembangan tanaman dan proses tanah-tanaman-atmosfer yang terhubung, seperti keseimbangan air dan nutrisi tanaman dan tanah, berdasarkan hubungan fisik dan biologis yang mendasarinya (Jones et al., 2003 ). Model berbasis proses melampaui korelasi empiris dengan mendefinisikan hubungan mekanistik dari proses mendasar yang mendorong pertumbuhan tanaman (Cuddington et al., 2013 ; Spitters, 1990 ). Hal ini harus dibedakan dari model yang murni digerakkan oleh data atau kecerdasan buatan dalam hal persyaratan data, penggabungan pengetahuan biofisik, dan penjelasan hasil, meskipun penelitian terkini menggabungkan keunggulan dari kedua pendekatan tersebut (Maestrini et al., 2022 ; Worrall et al., 2023 ; N. Zhang et al., 2023 ).
Sistem pendukung keputusan pertanian (DSS) menyusun model dan sumber data ini untuk mempelajari sistem pertanian dengan mengevaluasi kultivar, memperkirakan produksi dalam kondisi pertumbuhan yang berbeda, dan membandingkan alternatif manajemen. Model pertumbuhan tanaman dan DSS, seperti Sistem Pendukung Keputusan untuk Transfer Agroteknologi (DSSAT, Hoogenboom et al., 2019, 2024 ; Jones et al., 2003 ), sering disorot karena aplikasinya dalam pertanian presisi (Thorp et al., 2008 ). Mereka dapat berkontribusi untuk meningkatkan keberlanjutan praktik pertanian dengan memberikan informasi untuk meningkatkan output sambil mengurangi input (Aubert et al., 2012 ). Oleh karena itu, mengembangkan model berbasis proses untuk mensimulasikan pertumbuhan dan perkembangan rami industri dapat menjadi kontribusi yang signifikan untuk memajukan pemahaman kita tentang tanaman rami dan mengembangkan sistem tanam yang berkelanjutan.
Model-model sebelumnya yang mensimulasikan pertumbuhan dan perkembangan rami telah dikembangkan atau diadaptasi untuk kultivar serat dan benih terpilih yang tumbuh di lingkungan yang berbeda, yang dibangun di atas pendekatan pemodelan fisiologis yang berbeda. Dalam salah satu studi model rami pertama, Van der Werf et al. ( 1999 ) mensimulasikan potensi produksi rami di Eropa menggunakan model “Intersepsi dan Pemanfaatan Cahaya” (Spitters et al., 1989 ) dengan perkembangan yang bergantung pada suhu dan produksi dan pembagian bahan kering berdasarkan konsep intersepsi cahaya dan efisiensi penggunaan radiasi (RUE). Lisson et al. ( 2000 ) memodelkan kultivar rami serat yang berbeda di Australia dengan model “SIMulator Sistem Produksi Pertanian” (Keating et al., 2003 ). Fenologi dimodelkan menggunakan pendekatan derajat hari pertumbuhan dengan suhu dasar yang ditentukan secara empiris sebesar 1°C dan suhu optimum sebesar 29°C dan akumulasi biomassa didasarkan pada koefisien RUE yang dikalikan dengan radiasi yang dicegat. Radiasi yang dicegat dihitung menggunakan hukum Beer berdasarkan radiasi aktif fotosintesis (PAR) yang terjadi, koefisien kepunahan, dan indeks luas daun (LAI). Total LAI disimulasikan dengan menggabungkan proses individual untuk kemunculan daun, penuaan daun, dan perluasan daun sebagai fungsi waktu termal. Sebuah studi yang baru-baru ini diterbitkan oleh Žydelis et al. ( 2022 ) mengadaptasi model AgroC (Klosterhalfen et al., 2017 ) untuk rami, yang menggunakan SUCROS (Spitters et al., 1989 ) untuk komponen pertumbuhan tanaman. Fotosintesis dimodelkan menggunakan kurva respons cahaya untuk kedalaman tajuk yang berbeda dan diintegrasikan selama satu hari, dengan diferensiasi antara cahaya langsung dan cahaya difus dan karakteristik fotosintesis daun tunggal. Fotosintesis selanjutnya dikurangi dengan faktor stres kekeringan dan suhu, jika berlaku. Dalam pendekatan lain berdasarkan model “AquaCrop”, Steduto et al. ( 2009 ), Wimalasiri, Jahanshiri, Chimonyo, Azam-Ali, dkk. ( 2021 ), dan Wimalasiri, Jahanshiri, Chimonyo, Kuruppuarachchi, dkk. ( 2021 ) mengadaptasi model tebu untuk memprediksi serat dan biji rami. “AquaCrop” adalah model yang berfokus pada air. Oleh karena itu, fotosintesis tidak disertakan dan hanya diringkas dan diintegrasikan ke dalam satu koefisien produktivitas air. Perluasan luas daun secara tidak langsung disertakan dalam tutupan tanah yang tumbuh secara logistik, dikurangi oleh koefisien penuaan dan penuaan akhir musim. Berbeda dengan pendekatan yang disederhanakan ini, serangkaian eksperimen terperinci oleh Tang dkk. ( 2016, 2017 , 2018 ) menghasilkan kalibrasi model GECROS untuk rami (Komisi Eropa, 2017 ; Yin & van Laar, 2005). GECROS menggunakan model “Farquhar-von Caemmerer-Berry” yang secara biokimia lebih canggih (Farquhar et al., 1980 ) dengan fokus khusus pada penggandengan fotosintesis-transpirasi melalui konduktansi stomata dan interaksi karbon (C)–nitrogen (N) terperinci dengan luas daun. Berfokus secara eksklusif pada fenologi, Amaducci et al. ( 2008, 2012 ) mengembangkan model fenologi yang digerakkan oleh suhu untuk rami pasca-tumbuh dan aplikasi untuk produksi biomassa di seluruh Eropa. Terakhir, model rami serat lain oleh Baldini et al. ( 2020 ) secara langsung memprediksi hasil biji sebagai fungsi dari potensi hasil yang dikurangi oleh stres air atau suhu tanpa mensimulasikan fotosintesis atau proses fisiologis terperinci lainnya. Pendekatan pemodelan yang berbeda ini memberikan gambaran umum tentang aspek fisiologis utama tetapi juga skenario produksi dan pertanyaan penelitian yang berbeda untuk produksi rami. Kompleksitas atau kesederhanaan pendekatan pemodelan yang berbeda bukanlah penilaian kegunaan atau kualitas model, karena pilihan model yang tepat bergantung pada data yang tersedia, aplikasi yang dituju, dan jenis pengguna model, di antara faktor-faktor lainnya (KJ Boote et al., 1996 ).
Ide Inti
- Model pertumbuhan tanaman yang ada diadaptasi untuk produksi benih dan serat rami dengan data eksperimen Florida.
- Tanggal penanaman yang direkomendasikan di Florida utara adalah 1 Juni untuk produksi serat karena respons fotoperiode.
- Peningkatan produktivitas yang signifikan mungkin terjadi dengan panjang hari yang diperpanjang atau dengan kultivar panjang hari kritis yang lebih pendek.
Tinjauan kami terhadap aktivitas pemodelan sebelumnya pada rami industri menyimpulkan bahwa banyak model dan kumpulan data dasar tidak tersedia untuk umum, belum dikalibrasi dengan data dari Amerika Utara, dan tidak dibangun dengan fokus khusus pada evaluasi manajemen agronomi, termasuk aksesibilitas model untuk pengguna non-teknis. Oleh karena itu, untuk memajukan dan melengkapi aktivitas pemodelan sebelumnya, tujuan keseluruhan dari proyek penelitian ini adalah untuk mengembangkan model berbasis proses untuk rami berdasarkan Model Sistem Tanam (CSM) DSSAT untuk aplikasi lapangan. Tujuan khusus dari penelitian ini adalah untuk (1) memperoleh data eksperimen dari uji coba lapangan di Florida dan literatur untuk pengembangan model, (2) mengkalibrasi dan meningkatkan parameter genetik model dan persamaan fisiologis yang mendasarinya, dan (3) menunjukkan potensi aplikasi model melalui analisis musiman dari berbagai skenario produksi untuk dua lokasi di Florida dengan panjang hari, tanah, dan kondisi iklim yang berbeda.
2 BAHAN DAN METODE
2.1 Data Eksperimen
Uji coba rami agronomis dilakukan pada tahun 2021 dan 2022 di beberapa lokasi di Florida di bawah naungan Program Rami Industri Universitas Florida ( https://programs.ifas.ufl.edu/hemp ). Uji coba ini mencakup tanggal penanaman, tingkat pemupukan, dan kultivar yang berbeda. Pengembangan model awal didasarkan pada dua lokasi: Unit Penelitian dan Pendidikan Ilmu Tanaman (PSREU, https://plantscienceunit.ifas.ufl.edu ) dekat Citra di Florida utara-tengah (29.409° LU, −82.171° BB) dengan seri tanah pasir Arredondo dan Pusat Penelitian dan Pendidikan Florida Barat (WFREC, https://wfrec.ifas.ufl.edu ) dekat Jay di Florida barat laut (30.777° LU, 87.142° BB) dengan seri tanah lempung berpasir halus Dothan. Pengaturan eksperimen lengkap, termasuk deskripsi karakteristik tanah sebelum tanam, metodologi pengukuran, dan hasil untuk uji coba rami lain di lokasi ini yang tidak termasuk dalam proses pengembangan model, dapat ditemukan dalam publikasi masing-masing untuk lokasi PSREU (N. Kaur et al., 2024 ) dan lokasi WFREC (Nwosu et al., 2025 ). Sejauh memungkinkan, pengumpulan data eksperimen dilakukan sesuai dengan standar untuk pemodelan tanaman (KJ Boote et al., 2016 ; White et al., 2013 ) dan standar industri dan penelitian yang baru-baru ini ditetapkan untuk fisiologi dan morfologi rami industri (Brym et al., 2023 ).
Uji coba yang digunakan untuk adaptasi model mencakup kultivar serat IH Williams (IND Hemp, juga dikenal sebagai SS-Beta) dan kultivar serbaguna NWG 2730 (New West Genetics). Uji coba disusun dalam rancangan blok lengkap acak dengan enam tingkat pupuk nitrogen (N) (0, 56, 112, 168, 224, dan 280 kg N ha −1 ) dalam bentuk urea sebagai perlakuan. Plot berukuran panjang 7,62 m dan lebar 3,6 m dengan jarak baris tanam 0,2 m dan lorong 2 m atau lebih lebar di antara plot. Benih ditanam langsung dengan tingkat benih hidup 300.000–544.500 ha −1 pada tanggal yang berbeda di bulan Mei setiap tahun tergantung pada cuaca dan ketersediaan benih. Di lokasi PSREU, pupuk diberikan dalam aplikasi terpisah dengan 20% saat tanam, 30% pada 20 hari setelah tanam, dan 50% pada 40 hari setelah tanam. Lokasi WFREC memiliki aplikasi awal 56 kg N ha −1 pada 10 hari setelah tanam diikuti oleh jumlah N yang tersisa pada sekitar 35 hari setelah tanam menggunakan penebar tetes (GDY-6506TR-PPL, Gandy Company). Irigasi atas, dari sistem poros tengah di lokasi PSREU dan sistem pemindahan lateral di lokasi WFREC, diberikan dalam satu atau lebih dosis 13–19 mm selama 45 hari pertama setelah tanam untuk meningkatkan pembentukan tanaman dan menyerap N. Di lokasi WFREC, pengendalian gulma dilakukan sebelum tanam dengan 1,4 L ha −1 herbisida gulma berdaun lebar dan rumput (Dual Magnum, Syngenta) dan setelah tanam dengan pembudidaya atau penyiangan manual. Lokasi PSREU hanya melakukan pengendalian gulma pasca-tanam dengan kultivator atau penyiangan manual.
Di antara berbagai uji coba yang dilakukan di lokasi-lokasi ini, hanya eksperimen dengan kinerja terbaik dengan tekanan gulma minimal, tingkat perkecambahan yang dapat diterima, dan kinerja tanaman yang baik yang dipilih untuk adaptasi dan kalibrasi model. Sebanyak 15 kombinasi lokasi, tahun, kultivar, dan kombinasi pemupukan yang berbeda dipilih karena berkinerja baik dalam uji coba lapangan dan memiliki pengamatan terbanyak, dan dijelaskan lebih rinci dalam Bagian “ 2.4 Pengaturan dan Input Simulasi” dan Tabel 2. Namun, dibandingkan dengan lokasi lain dan khususnya lokasi di Eropa atau Cina dengan riwayat budidaya yang lebih panjang, produktivitas biomassa kering total umumnya lebih rendah karena fase pertumbuhan vegetatif yang lebih pendek dan pembungaan awal serta tantangan umum dengan kualitas benih dan pembentukan tanaman. Pengukuran untuk biomassa total, partisi jaringan, jumlah simpul pada batang utama, tinggi tanaman, dan lebar tanaman maksimum diperoleh beberapa kali sepanjang musim dengan tiga atau lebih pengulangan per perlakuan. Untuk setiap pengukuran biomassa, tanaman dari area representatif sekitar 0,25 m × 0,25 m pada tahap pertumbuhan awal yang meningkat menjadi 1,0 m × 1,0 m pada tahap pertumbuhan selanjutnya dengan peningkatan ukuran tanaman dipanen dan dianalisis. Tinggi tanaman, lebar, dan jumlah buku diukur pada tanaman yang disingkirkan dan pada tanggal tambahan sepanjang musim pada tanaman yang berdiri tanpa memanennya. Selain itu, pengamatan fenologi dicatat untuk beberapa percobaan, termasuk kemunculan, awal pembungaan (bunga terminal pertama), pembungaan penuh (50% tanaman jantan pembungaan terminal), awal pembentukan biji, dan saat sebagian besar biji matang. Pembungaan terminal didefinisikan sebagai kelompok bunga pertama yang ditentukan di puncak pucuk mengikuti Brym et al. ( 2023 ). Pengamatan yang hilang untuk pembungaan terminal diperkirakan 10 hari sebelum 50% tanaman jantan berbunga berdasarkan rangkaian waktu pengamatan fenologi di lokasi lain. Jarak tempuh membatasi jumlah kunjungan lapangan yang memungkinkan, yang menyebabkan jumlah pengukuran yang bervariasi untuk beberapa percobaan. Tingkat variabilitas yang tinggi ditemukan antara blok-blok dengan perlakuan yang sama, kemungkinan karena variasi di dalam lahan dan tekanan gulma di beberapa plot. Selain eksperimen lapangan khusus yang dijelaskan, pengetahuan yang lebih umum tentang fisiologi rami, fenologi, dan respons manajemen yang diperoleh dari lokasi yang tersisa dari Program Rami Industri Universitas Florida juga dipertimbangkan selama pengembangan model dan proses adaptasi.
2.2 Data cuaca
Data cuaca untuk dua lokasi eksperimen diperoleh dari Florida Automated Weather Network (FAWN, Lusher et al., 2008 ). Stasiun “Citra” (29.410° N 82.173° W) dipasang pada tahun 2003 dan berlokasi dekat uji coba lapangan PSREU. Stasiun “Jay” (30.775° N 87.141° W) dipasang pada tahun 2003 dan berlokasi dekat uji coba lapangan WFREC. Kedua stasiun menyediakan suhu udara minimum dan maksimum harian, total presipitasi harian, total radiasi matahari harian, kecepatan angin, dan kelembaban relatif, selain pengukuran yang lebih rinci (Tabel 1 ). Kedua lokasi lapangan dicirikan oleh iklim subtropis lembab dengan musim hujan dari Mei hingga Oktober. Panjang hari, yang relevan untuk simulasi fotoperiode dan fenologi, dihitung berdasarkan deklinasi matahari, lintang, dan bujur stasiun. Untuk mengevaluasi simulasi tanggal pembungaan untuk lokasi di luar Florida, data cuaca yang diperoleh dari satelit dari NASA-Power (NASA, 2018 ) diperoleh untuk Urbana, IL (40.114° LU, 88.234° BB); Hyde, MI (45.734° LU, 87.203° BB); dan Manhattan, KS (39.184° LU, 96.572° BB). Lokasi tersebut memiliki pengamatan fenologi di Midwestern Hemp Database (Alberti, 2021 ; University of Illinois, 2024 ) untuk kultivar yang sama tetapi pada garis lintang yang berbeda yang membuatnya cocok untuk evaluasi simulasi fenologi.
| Lokasi | Periode waktu | Suhu rata-rata (°C) | Titik embun rata-rata (°C) | Kelembaban relatif rata-rata (%) | Curah hujan bulanan kumulatif (mm) | Rata-rata total radiasi matahari harian (MJ m − 2 ) | Rata-rata kecepatan angin harian (km h − 1 ) |
|---|---|---|---|---|---|---|---|
| PSREU | Mei 2021 | 23.9 | 17.5 | 71 | 47 | 21.1 | 8.2 |
| Juni 2021 | 25.8 | 22.9 | 86 | 334 | 15.0 | 7.2 | |
| Juli 2021 | 26.6 | 24.0 | 87 | 156 | 16.0 | 6.6 | |
| Agustus 2021 | 27.1 | 24.5 | 87 | 168 | 15.4 | 6.8 | |
| September 2021 | 25.3 | 22.5 | 86 | 167 | 14.4 | 5.3 | |
| Mei 2022 | 24.7 | 20.0 | 78 | 109 | 20.1 | 7.2 | |
| Juni 2022 | 27.4 | 23.1 | 80 | 60 | 20.3 | 7.2 | |
| Juli 2022 | 27.2 | 24.0 | 85 | 137 | 18.1 | 6 | |
| Agustus 2022 | 26.9 | 23.9 | 86 | 256 | 16.4 | 5.1 | |
| September 2022 | 25.3 | 22.7 | 87 | 122 | 13.3 | 6.4 | |
| Bahasa Inggris WFREC | Mei 2021 | 22.2 | 17.1 | 76 | 198 | 22.1 | 10.6 |
| Juni 2021 | 25.4 | 22.7 | 86 | 321 | 19.1 | 9.2 | |
| Juli 2021 | 26.0 | 23.9 | 90 | 295 | 18.3 | 6.1 | |
| Agustus 2021 | 26.3 | 23.9 | 88 | 300 | 18.2 | 7.4 | |
| September 2021 | 23.7 | 21.2 | 88 | 187 | 16.0 | 6.6 | |
| Mei 2022 | 23.7 | 19.7 | 81 | 253 | 21.0 | 9.3 | |
| Juni 2022 | 26.9 | 22.9 | 81 | 106 | 22.1 | 7.2 | |
| Juli 2022 | 26.0 | 24.1 | 90 | 262 | 17.8 | 6 | |
| Agustus 2022 | 25.5 | 23.6 | 90 | 197 | 16.7 | 6 | |
| September 2022 | 23.9 | 19.4 | 79 | 10 | 18.7 | 7.7 |
Catatan : Lokasi PSREU adalah Plant Science Research and Education Unit di Citra, FL. Lokasi WFREC adalah West Florida Research and Education Center di Jay, FL. Kecepatan angin rata-rata diambil pada ketinggian 10 m di atas tanah; semua pengukuran lainnya diambil pada ketinggian 2 m di atas tanah.
2.3 Perangkat lunak, kode sumber, dan statistik
Simulasi tanaman dilakukan dengan model CSM (Jones et al., 2003 ) dalam perangkat lunak pemodelan DSSAT 4.8.2 (Hoogenboom et al., 2019 , 2024 ) yang tersedia di http://www.dssat.net . Kode sumber tersedia melalui repositori GitHub di https://github.com/DSSAT/dssat-csm-os . DSSAT mencakup berbagai macam model dan kelompok model, dengan infrastruktur data menyeluruh, antarmuka pengguna grafis, dan fungsi pendukung. CROPGRO adalah satu kelompok model spesifik dalam CSM dan merupakan kerangka kerja untuk pengembangan model rami. Parameter model yang diubah selama pengembangan model untuk rami akan disajikan di Bagian “ 3.1 Proses Pengembangan Model.” Set lengkap parameter dan file yang diperlukan untuk CSM–CROPGRO-Hemp, yang kemudian disebut sebagai CROPGRO-Hemp, disediakan sebagai Informasi Pendukung 1 dan akan diimplementasikan dalam versi DSSAT mendatang untuk rilis publik. Analisis statistik dan visualisasi hasil dilakukan di R-Studio 2023.06.2 (Tim RStudio, 2020 ) dan R 3.4.2 (Tim Inti R, 2021 ). Paket “DSSAT” digunakan untuk menjalankan simulasi DSSAT secara otomatis di antarmuka R, “ggplot2” untuk memplot data simulasi dan observasi, dan “tidyverse,” “dplyr,” “hydroGOF,” dan “ggpubr” untuk analisis data. Skema dan visualisasi tambahan dibuat di LaTeX menggunakan paket “TikZ.” Rekomendasi umum yang diberikan oleh J.-Y. Yang et al. ( 2014 ) diikuti untuk analisis keseluruhan. Indeks Kesepakatan Model Wilmott ( d , [Persamaan 1 ]) digunakan untuk menilai kesepakatan antara prediksi model dan data observasi dan, dengan demikian, kinerja model umum (Willmott et al., 2012 ). Ini adalah indeks tanpa dimensi yang terkait dengan indeks efisiensi Nash–Sutcliffe dengan nilai berkisar antara 0,0 dan 1,0, dengan nilai yang mendekati 1 menunjukkan kinerja model yang lebih baik.

2.4 Pengaturan dan input simulasi
Input model tanaman dan file simulasi dibuat untuk setiap percobaan individual agar menyerupai uji coba lapangan dan mensimulasikan pertumbuhan dan perkembangan di bawah input N nol dan aplikasi pupuk N tinggi (Tabel 2 ). Tingkat maksimum 280 kg N ha −1 yang diterapkan dalam percobaan berada di ujung atas, jika tidak melampaui, kisaran pemupukan N yang direkomendasikan yang diidentifikasi dalam literatur (N. Kaur et al., 2023 ). Perlakuan ini dipilih untuk pengembangan model untuk mensimulasikan skenario yang dekat dengan produksi potensial yang hanya dibatasi oleh kondisi cuaca. Dalam percobaan yang dipilih, tingkat N yang lebih rendah dipilih untuk pengaturan simulasi karena lebih banyak pengamatan dalam musim tersedia untuk perlakuan yang lebih rendah atau karena tingkat N yang tinggi berkinerja lebih buruk daripada tingkat N sedang. Efek eksternal yang merugikan atau ketidakteraturan yang mendasarinya dalam kondisi lapangan lokal akan menjadi tantangan untuk diukur dan diwakili secara memadai dalam versi model awal. Karena tingkat kemunculan benih yang bervariasi dan penjarangan selama minggu-minggu pertama pembentukan tanaman, tegakan tanaman yang diamati secara signifikan di bawah kepadatan tanaman yang ditargetkan. Karena penjarangan sendiri belum disertakan dalam kerangka model CROPGRO, kerapatan penanaman untuk tujuan pemodelan didasarkan pada jumlah tegakan yang diperoleh pada 4 minggu setelah kemunculan pertama, dengan asumsi bahwa tanaman ini akan bertahan hidup hingga akhir musim. Simulasi dinamika kalium (K) dan fosfor (P) dan simbiosis bakteri pengikat N dinonaktifkan dalam pengaturan simulasi karena kurangnya data yang sesuai untuk kalibrasi dan karena tidak berlaku untuk rami sebagai tanaman non-kacang-kacangan, meskipun kerangka DSSAT secara umum mendukung kemampuan ini. Diaktifkan oleh struktur modular DSSAT dan CROPGRO, beberapa submodul tersedia untuk mensimulasikan proses biofisika dan biokimia yang sama berdasarkan prinsip atau asumsi ilmiah yang berbeda. Modul “CENTURY” (Gijsman et al., 2002 ) dipilih untuk bahan organik tanah, “Ritchie Water Balance” (Ritchie, 1998 ) untuk hidrologi, “Soil Conservation Series” untuk infiltrasi, “Ritchie-Ceres” untuk penguapan tanah, “Modified Soil Profile” untuk distribusi horizon tanah, dan “Priestley-Taylor” untuk evapotranspirasi potensial (Hoogenboom et al., 2019 ). Pemilihan masing-masing submodul didasarkan pada CROPGRO-Soybean ( Glycine max) dan dianggap layak mengingat data eksperimen yang tersedia dan hasil pemodelan yang diperoleh. Pertanyaan penelitian, lokasi, dan data eksperimen yang berbeda di masa mendatang mungkin memerlukan evaluasi modul alternatif. Untuk mengevaluasi simulasi tanggal pembungaan untuk lokasi di luar Florida, simulasi yang disederhanakan disiapkan dengan dinamika air dan N yang dinonaktifkan serta profil tanah standar dan informasi pengelolaan, karena hanya simulasi fenologi yang menarik.
| Simulasi | |||||||
|---|---|---|---|---|---|---|---|
| TIDAK. | Kode | Lokasi | Tahun | Kultivar | Pupuk (kg N ha − 1 ) | Tanggal Penanaman | Kepadatan (tanaman m − 2 ) |
| 1 | UFCI2101 | PSREU | Tahun 2021 | IH Williams | 280 | 5 Mei 2021 | 53.8 |
| 2 | UFCI2101 | PSREU | Tahun 2021 | Kelompok Kerja Nasional 2730 | angka 0 | 5 Mei 2021 | 64.0 |
| 3 | UFCI2101 | PSREU | Tahun 2021 | Kelompok Kerja Nasional 2730 | 280 | 5 Mei 2021 | 60.5 |
| 4 | UFCI2201 | PSREU | Tahun 2022 | IH Williams | angka 0 | 4 Mei 2022 | 50.7 |
| 5 | UFCI2201 | PSREU | Tahun 2022 | IH Williams | 280 | 4 Mei 2022 | 40.8 |
| 6 | UFCI2201 | PSREU | Tahun 2022 | Kelompok Kerja Nasional 2730 | angka 0 | 4 Mei 2022 | 30.5 |
| 7 | UFCI2201 | PSREU | Tahun 2022 | Kelompok Kerja Nasional 2730 | 168 | 4 Mei 2022 | 40.5 |
| 8 | UFJA2101 | Bahasa Inggris WFREC | Tahun 2021 | IH Williams | angka 0 | 26 Mei 2021 | 66.8 |
| 9 | UFJA2101 | Bahasa Inggris WFREC | Tahun 2021 | IH Williams | 224 | 26 Mei 2021 | 68.6 |
| 10 | UFJA2101 | Bahasa Inggris WFREC | Tahun 2021 | Kelompok Kerja Nasional 2730 | angka 0 | 26 Mei 2021 | 71.5 |
| 11 | UFJA2101 | Bahasa Inggris WFREC | Tahun 2021 | Kelompok Kerja Nasional 2730 | 112 | 26 Mei 2021 | 71.5 |
| 12 | UFJA2201 | Bahasa Inggris WFREC | Tahun 2022 | IH Williams | angka 0 | 13 Mei 2022 | 99.9 |
| 13 | UFJA2201 | Bahasa Inggris WFREC | Tahun 2022 | IH Williams | 280 | 13 Mei 2022 | 71.9 |
| 14 | UFJA2201 | Bahasa Inggris WFREC | Tahun 2022 | Kelompok Kerja Nasional 2730 | angka 0 | 13 Mei 2022 | 69.8 |
| 15 | UFJA2201 | Bahasa Inggris WFREC | Tahun 2022 | Kelompok Kerja Nasional 2730 | 280 | 13 Mei 2022 | 82.8 |
Catatan : Lokasi PSREU adalah Plant Science Research and Education Unit di Citra, FL. Lokasi WFREC adalah West Florida Research and Education Center di Jay, FL. Kepadatan penanaman untuk pemodelan didasarkan pada jumlah tegakan pada 4 minggu setelah kemunculan. Kultivar IH Williams juga dikenal sebagai SS-Beta. Tidak ada eksperimen 0 kg N ha − 1 yang dilakukan untuk IH Williams pada tahun 2021 di lokasi PSREU.
2.5 Data tanah
Informasi profil tanah (Tabel 3 ) diperoleh melalui pengukuran dan diestimasi berdasarkan perbandingan dengan profil tanah dari studi pemodelan tanaman sebelumnya dan deskripsi seri tanah untuk lempung berpasir halus Dothan di WFREC (KJ Boote et al., 2021 ; USDA NRCS, 2017 ) dan untuk pasir Arredondo di PSREU (de Souza Nóia Júnior et al., 2022 ; USDA NRCS, 2018 ). Kumpulan karbon organik tanah yang stabil ditentukan secara manual selama proses kalibrasi model untuk mencapai respons pertumbuhan biomassa tanaman yang diinginkan terhadap perlakuan pupuk N nol. Langkah ini diperlukan saat menggunakan modul CENTURY di DSSAT untuk mensimulasikan proses tanah–tanaman–nitrogen secara memadai, khususnya dalam eksperimen tanpa atau dengan masukan N rendah (Gijsman et al., 2002 ). Parameter air tanah “SLLL,” “SDUL,” dan “SSAT” menentukan kemampuan menahan air dan dihitung berdasarkan tekstur tanah. Jika tidak ada pengukuran, air tanah awal diperkirakan sebesar 75% dari air yang tersedia dan kandungan N tanah sebesar 50 kg N ha −1 yang disebarkan secara merata di semua lapisan tanah pada tanggal 1 Januari setiap musim untuk memungkinkan modul CENTURY diinisialisasi hingga penanaman. Residu tanaman rami dan bahiagrass ( Paspalum notatum ) sebelumnya diperkirakan sebesar 2000 kg ha −1 dengan konsentrasi N sebesar 1%.
| Tekstur tanah | Air tanah | Nitrogen tanah | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Kedalaman (m) | Tanah liat (%) | Lumpur (%) | Berat jenis (g cm – 3 ) | SLLL (cm 3 cm − 3 ) | SDUL (cm 3 cm − 3 ) | SSAT (cm 3 cm − 3 ) | SH2O ( cm3cm − 3 ) *** | Tingkat Kemiringan (%) | SASC (%) | SNH4 (unsur g N Mg − 1 )*** | SNO3 (g unsur N Mg − 1 )*** |
| PSREU | |||||||||||
| 0,05 | 2.4 | 7.1 | 1.42 | 0,070 | 0.142 | 0.493 | 0.124 | 0,75 | 0,56/0,64* | 0.2 | 1.9 |
| 0,15 | 2.1 | 6.3 | 1.45 | 0,024 | 0.161 | 0.487 | 0.127 | 0,75 | 0,56/0,64* | 0.2 | 1.9 |
| 0.30 | 2.3 | 4.1 | 1.59 | 0,016 | 0.104 | 0.407 | 0,082 | 0.34 | 0,26/0,29* | 0.2 | 1.9 |
| 0,50 | 2.2 | 4.0 | 1.59 | 0,014 tahun | 0.106 | 0.405 | 0,083 tahun | 0.14 | 0,11/0,12* | 0.2 | 1.9 |
| 0,90 | 2.4 | 3.3 | 1.60 | 0,011 | 0.113 | 0.408 | 0,087 tahun | 0,09 | 0,07/0,08* | 0.2 | 1.9 |
| 1.20 | 1.9 | 3.3 | 1.60 | 0,021 | 0,08 | 0,385 | 0,065 tahun | 0,07 | 0,05/0,06* | 0.2 | 1.9 |
| 1.50 | 1.9 | 3.3 | 1.63 | 0,021 | 0,08 | 0,385 | 0,065 tahun | 0,07 | 0,05/0,06* | 0.2 | 1.9 |
| Bahasa Inggris WFREC | |||||||||||
| 0,05 | 15 | 15.5 | 1.6 | 0.103 | 0.218 | 0.406 | 0.189 | 0,89 | 0,53/0,49** | 0.2 | 1.9 |
| 0,15 | 15 | 15.5 | 1.6 | 0.103 | 0.218 | 0.406 | 0.189 | 0,89 | 0,53/0,49** | 0.2 | 1.9 |
| 0.30 | 20 | 12.5 | 1.6 | 0,125 | 0.239 | 0.413 | 0.210 | 0.32 | 0,19/0,18** | 0.2 | 1.9 |
| 0,60 | 20 | 12 | 1.6 | 0,135 | 0.251 | 0.415 | 0.222 | 0.10 | 0,06/0,06** | 0.2 | 1.9 |
| 0,90 | 20 | 11 | 1.6 | 0,147 tahun | 0.261 | 0.417 | 0.233 | 0,04 | 0,02/0,02** | 0.2 | 1.9 |
| 1.20 | 20 | 10 | 1.6 | 0,147 tahun | 0.261 | 0.417 | 0.233 | 0,04 | 0,02/0,02** | 0.2 | 1.9 |
| 1.50 | 20 | 7 | 1.6 | 0,125 | 0.236 | 0.412 | 0.208 | 0,02 | 0,01/0,01** | 0.2 | 1.9 |
Catatan : Lokasi PSREU adalah Plant Science Research and Education Unit di Citra, FL. Lokasi WFREC adalah West Florida Research and Education Center di Jay, FL. BD = bulk density. Batas bawah tanah untuk kadar air tanah yang dapat diekstraksi tanaman (SLLL), batas atas yang dikeringkan (SDUL) dan saturasi (SSAT) menentukan kemampuan tanah menahan air. Karbon organik tanah yang stabil (SASC) dinyatakan dalam satuan absolut, tetapi sebagai persentase karbon organik tanah (SLOC), yang berjumlah 75% (*) pada tahun 2021, 80% pada tahun 2022 di lokasi PSREU 60% (**) pada tahun 2021, 55% pada tahun 2022 di lokasi WFREC. Air tanah (SH2O), amonium tanah (SNH4), dan nitrat tanah (SNO3) awal (***) diperkirakan sebesar 75% air tersedia dan 50 kg N ha − 1 pada tanggal 1 Januari setiap musim.
2.6 Proses adaptasi dan kalibrasi model
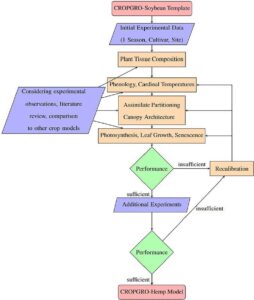
Model yang ada untuk kedelai dan kapas dari CSM-CROPGRO digunakan sebagai templat, karena mereka adalah tanaman yang dipelajari dengan baik dengan pengembangan model yang luas, aplikasi, dan data eksperimen pendukung untuk parameter model (K. Boote et al., 1997 ; Hoogenboom et al., 1992 ; Li et al., 2019 ; Thorp et al., 2014 ). Parameter CROPGRO untuk kedelai dan kapas, oleh karena itu, berfungsi sebagai nilai referensi yang dibandingkan dengan rami. Lebih jauh, baik aspek biji-bijian maupun serat dari produksi rami adalah aspek yang relevan dalam studi ini, yang memiliki kesamaan dengan sistem produksi kedelai dan kapas. CROPGRO adalah templat pemodelan generik dalam DSSAT, yang awalnya dikembangkan untuk kacang-kacangan biji-bijian dengan perhitungan fotosintesis tingkat daun per jam yang digabungkan dengan intersepsi cahaya tipe pagar tanaman (K. Boote & Pickering, 1994 ). Selain itu, CROPGRO juga memuat keseimbangan tanah-tanaman-nitrogen dan air serta proses pertumbuhan dan perkembangan fotosintesis, vegetatif, dan reproduktif (KJ Boote, Jones, & Hoogenboom, 1998 ; KJ Boote, Jones, Hoogenboom, et al., 1998 ). Parameter fisiologis dan genetik dalam CROPGRO menggambarkan sifat kompleks tanaman pangan, yang dipisahkan menjadi parameter spesies, ekotipe, dan kultivar. Organisasi hierarkis bertujuan untuk menyediakan landasan universal bagi tanaman pangan (parameter spesies), yang kemudian disesuaikan lebih lanjut untuk kelompok ekotipe dan kultivar individual yang berbeda. Templat dan parameter untuk file spesies, ekotipe, dan kultivar disesuaikan mengikuti contoh adaptasi terbaru model CROPGRO untuk guar ( Cyamopsis tetragonoloba ; KJ Boote et al., 2023 ), carinata ( Brassica carinata ; KJ Boote et al., 2021 ), stroberi ( Fragaria × ananassa ; Hopf et al., 2022 ), safflower ( Carthamus tinctorius ; Singh et al., 2016 ), dan alfalfa ( Medicago sativa; Malik et al., 2018 ). Keseluruhan proses didominasi oleh manual, dengan penyesuaian parameter yang berulang dan perbandingan antara keluaran model simulasi dan data yang diamati (Gambar 1 ). Sementara beberapa parameter didefinisikan dengan jelas melalui bukti eksperimental atau tinjauan pustaka, parameter lain diperkirakan dari berbagai nilai yang layak di seluruh spesies tanaman CSM-CROPGRO lainnya. Alat optimasi GLUE dan GENCALC (He et al., 2010 ; Hunt et al., 1993 ; Jones et al., 2011 ) yang disertakan dalam DSSAT digunakan secara terbatas untuk kalibrasi parameter kultivar untuk fenologi.
2.7 Analisis musiman
Untuk menunjukkan aplikasi potensial dari model rami baru, analisis musiman (Thornton & Hoogenboom, 1994 ) dilakukan dengan menggunakan data cuaca historis selama 21 tahun. Data cuaca harian dari stasiun cuaca FAWN di Jay dan Citra di Florida digunakan untuk mensimulasikan 21 musim tanam antara tahun 2003 dan 2023 dan memperkirakan biomassa dan produksi biji-bijian. Sejumlah kecil pengamatan cuaca yang keliru, yaitu suhu beku pada bulan Juni atau nilai yang hilang, digantikan dengan mengambil rata-rata nilai masing-masing dari hari sebelumnya dan berikutnya. Sebagai faktor tambahan, variasi tanggal tanam dievaluasi dengan interval 15 hari dari 1 Maret hingga 30 Juni setiap tahun untuk mempelajari dampak tanggal tanam yang lebih awal atau lebih lambat. Kepadatan tanaman ditetapkan sebesar 80 tanaman m −2 untuk IH Williams dan NWG 2730, yang berada di kisaran atas kepadatan tanaman mapan yang diamati 4 minggu setelah penaburan. Pengelolaan tanaman identik dengan percobaan lapangan yang dijelaskan sebelumnya, dengan irigasi atas setelah penanaman dan pemupukan 280 kg N ha −1 yang diberikan sebagian pada saat penanaman, diikuti dengan pembagian pada hari-hari tertentu setelah penanaman, disesuaikan dengan tanggal penanaman yang berbeda. Profil tanah diinisialisasi dengan 75% air tersedia dan 50 kg N ha −1pada tanggal 1 Januari setiap musim dan kumpulan karbon organik tanah yang stabil ditetapkan sebesar 80% dari karbon organik tanah yang tersedia di PSREU dan sebesar 60% di WFREC. Tanggal panen untuk tujuan serat ditetapkan terjadi pada simulasi pembentukan benih pertama, yaitu 10 hari fototermal (PD) atau sekitar 2 minggu dalam hari kalender setelah “bunga terminal pertama” atau sekitar tanggal “pembungaan penuh,” sekitar 35–60 hari setelah penanaman di seluruh tanggal penanaman yang berbeda. Ini dicapai dalam model dengan membuat kultivar hipotetis dengan koefisien genetik untuk waktu dari benih hingga kematangan fisiologis (SD-PM) dan kematangan panen (R7–R8) masing-masing ditetapkan sebesar 0,1 PD. Tanaman serat kemudian mencapai kematangan panen segera setelah dimulainya pembentukan benih. Tanggal panen untuk tujuan biji-bijian ditetapkan berdasarkan pengamatan di lokasi PSREU dengan tanggal puncak pematangan benih sebelum benih pecah berlebihan atau penuaan tanaman. Untuk memungkinkan benih yang terbentuk terlambat untuk matang dalam model, periode tambahan setelah hari simulasi kematangan fisiologis benih pertama hingga panen ditetapkan sebagai sekitar 1 bulan untuk kultivar IH Williams dan sekitar 2 minggu untuk NWG 2730, menghasilkan panen sekitar 85–140 hari kalender setelah penanaman. Untuk jenis analisis kedua, dua perlakuan dengan tiga level masing-masing dibuat untuk menganalisis dampak perubahan hipotetis pada produksi rami di lokasi ini. Untuk mensimulasikan potensi kemajuan pemuliaan, kami menetapkan kultivar hipotetis yang identik dengan IH Williams tetapi dengan panjang hari kritis (CSDL) yang lebih rendah sebesar 0,25, 0,50, atau 1,00 jam. CSDL menetapkan panjang periode cahaya dalam siklus 24 jam yang diperlukan untuk menghambat atau menunda pembungaan pada tanaman hari pendek seperti rami. Sebagai respon manajemen, perlakuan kedua memperpanjang total panjang hari sebanyak 0,5, 1,0, atau 2,0 jam dibandingkan dengan panjang hari yang diamati untuk setiap lokasi, yang mewakili penggunaan pencahayaan tambahan berintensitas rendah di lingkungan hari pendek atau produksi di lintang lebih utara dengan iklim serupa tetapi panjang hari lebih panjang.
3 HASIL DAN PEMBAHASAN
3.1 Proses pengembangan model
Berdasarkan proses adaptasi model dan kalibrasi parameter yang dijelaskan sebelumnya, beberapa kelompok parameter model disesuaikan dalam proses berulang. Pada bagian selanjutnya, hubungan khusus antara fisiologi rami, proses model yang sesuai, dan justifikasi untuk pemilihan parameter model akan disajikan sebelum membahas kinerja model yang telah difinalisasi.
3.1.1 Komposisi jaringan tumbuhan
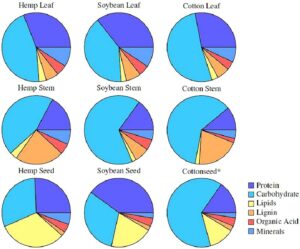
Komposisi jaringan tanaman menentukan jumlah asimilat yang dibutuhkan untuk membentuk berbagai jenis jaringan dan dikaitkan dengan keseimbangan karbon dan N secara keseluruhan dari model CROPGRO. Komposisi makromolekul rata-rata dari berbagai jaringan tanaman rami diperoleh dari literatur (Audu et al., 2014 ; Kleinhenz et al., 2020 ; Xu et al., 2022 ), dengan serat sebagai lignin dan abu sebagai kandungan mineral yang sejalan dengan terminologi CROPGRO. Dalam literatur, komposisi lipid (minyak) jaringan rami yang dilaporkan mencapai 20% untuk jaringan daun dan 8% untuk jaringan batang. Ini tinggi untuk tanaman serat dan kemungkinan terjadi karena pengukuran diperoleh dari bunga rami yang kaya kanabinoid dengan kelenjar resin. Menggunakan konsentrasi lipid yang tinggi akan membiaskan simulasi; oleh karena itu, untuk CROPGRO-Hemp, kami mengurangi komposisi lipid daun dan batang menjadi 3,5% yang mendukung kandungan karbohidrat dan serat yang lebih tinggi. Informasi tentang kandungan asam organik dari berbagai jaringan rami dan komposisi umum akar rami tidak ditemukan dan, oleh karena itu, diasumsikan sama dengan kedelai (Gambar 2 ). Khusus untuk komposisi protein, batas atas dan bawah konsentrasi N jaringan ditentukan untuk mensimulasikan pertumbuhan jaringan di bawah pertumbuhan yang dibatasi atau tidak terbatas N. Oleh karena itu, komposisi protein jaringan yang disimulasikan pada waktu tertentu bergantung pada dinamika sumber-penyerap secara keseluruhan, seperti ketersediaan N serta fase fenologi yang mendorong pertumbuhan, reproduksi, penuaan, dan mobilisasi serta realokasi N dari jaringan yang lebih tua ke jaringan yang lebih baru. Dinamika N CROPGRO dijelaskan secara lebih rinci dalam KJ Boote et al .
3.1.2 Arsitektur kanopi
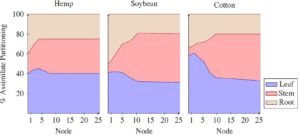
Arsitektur kanopi di CROPGRO menentukan intersepsi cahaya pagar tanaman per jam dan fotosintesis tingkat daun karena kanopi yang lebih tinggi dan lebih lebar umumnya mengarah pada penutupan kanopi yang lebih lengkap dan penyerapan cahaya. Untuk rami, kami mendefinisikan 25 simpul di sepanjang batang utama, setara dengan 50 daun dalam susunan daun yang berlawanan dengan dua daun per simpul, sebagai pola khas berdasarkan pengamatan eksperimental. Jumlah simpul yang sebenarnya diproduksi akan bergantung pada efek panjang hari untuk menginduksi pembungaan lebih awal atau lebih lambat. Panjang ruas dan lebar kanopi per simpul disesuaikan untuk mengikuti pola pertumbuhan yang diamati dalam tinggi dan lebar. Misalnya, ruas lebih panjang dari ruas 7 hingga 13 agar sesuai dengan perkembangan kanopi cepat yang diamati selama fase pertumbuhan vegetatif. Fase ini didahului oleh pertumbuhan tinggi yang lebih lambat selama pembentukan tanaman dan diikuti oleh pertumbuhan tinggi yang lebih lambat setelah pembungaan menjelang akhir siklus pertumbuhan selama pembentukan dan pematangan benih (Tabel 4 ). Selain itu, koefisien genetik “RHGHT” mendefinisikan tinggi setiap kultivar relatif terhadap tinggi standar dan ditetapkan pada 1,2 untuk IH Williams dan pada 0,8 untuk NWG 2730 untuk mensimulasikan kanopi IH Williams yang lebih tinggi secara keseluruhan. Konsep “V-Stage” (jumlah node) dalam CROPGRO mendorong koefisien partisi untuk pertumbuhan daun, batang, dan akar dalam kaitannya dengan jumlah node pada batang utama. Selama simulasi, fungsi pencarian dengan interpolasi linier mendefinisikan tingkat partisi asimilasi saat ini berdasarkan V-Stage saat ini. Mekanisme ini, dikombinasikan dengan dinamika sumber-penyerap, keterbatasan stres, penuaan, serta keberadaan organ reproduksi (benih) yang diprioritaskan untuk asimilat, mendorong partisi asimilat yang bervariasi dan dinamika pertumbuhan organ sepanjang siklus pertumbuhan (Gambar 3 ). Dibandingkan dengan kedelai dan kapas, parameter partisi asimilat untuk rami sedikit disesuaikan untuk mencapai pertumbuhan daun awal musim dan penutupan kanopi yang diamati. CROPGRO membatasi lebar tajuk maksimum pada jarak baris, dengan asumsi tutupan tajuk penuh tercapai setelah tanaman mencapai lebar yang sama dengan lebar baris. Ini tidak memperhitungkan tanaman yang mengembangkan tajuk yang lebih lebar dan tumbuh ke baris tetangga seperti yang diamati dalam uji coba rami. Ini dapat dievaluasi dengan meningkatkan jarak baris dalam pengaturan simulasi menjadi lebih besar dari jarak baris sebenarnya, tetapi ternyata ini hanya memiliki efek minimal pada pola pertumbuhan keseluruhan dan hasil simulasi.
| Proses | Koefisien genetik | Rami | Kedelai |
|---|---|---|---|
| Asimilasi partisi | Berat relatif benih yang menjadi tanaman awal (fraksi, WTFSD) | 0,60 | 0,55 |
| Rasio berat tangkai daun terhadap berat daun (PORPT) | 0,07 | 0,58 | |
| Fraksi asimilat dibagi ke pertumbuhan batang harian setelah tahap V maksimum (g[batang] g[sayur] − 1 , FRSTMF) | 0,45 | 0,55 | |
| Fraksi asimilat dibagi ke dalam pertumbuhan daun harian setelah maksimum
Tahapan V (g[daun] g[sayur] − 1 , FRLFF) |
0,35 | 0.24 | |
| Pembagian maksimum ke daun (g[daun] g[sayuran] − 1 , FRLFMX) | 0.8 | 0.7 | |
| Pertumbuhan kanopi | Jumlah node spesifik spesies []* serta panjang ruas dan lebar tajuk per node dalam (m) | [0,0] * 0,015/0,030 | [0,0] 0,034/0,034 |
| [2.0] * 0,035/0,040 | [1.0] 0,056/0,054 | ||
| [4.0] * 0,070/0,050 | [4.0] 0,065/0,064 | ||
| [7.0] * 0,130/0,060 | [6.0] 0,067/0,065 | ||
| [10.0] * 0,140/0,070 | [8.0] 0,069/0,066 | ||
| [13.0] * 0,160/0,060 | [10.0] 0,066/0,062 | ||
| [16.0] * 0,060/0,060 | [14.0] 0,060/0,057 | ||
| [19.0] * 0,030/0,060 | [16.0] 0,050/0,044 | ||
| [25.0] * 0,020/0,040 | [20.0] 0,034/0,025 | ||
| [40.0] * 0,010/0,020 | [40.0] 0,006/0,001 |
3.1.3 Suhu kardinal
Suhu kardinal menentukan sensitivitas spesies terhadap suhu selama fase pertumbuhan yang berbeda atau untuk proses tertentu dan berlaku untuk semua kultivar. Mereka khususnya relevan untuk simulasi fenologi, yang akan dibahas lebih rinci dalam Subbagian “ 3.1.6 Fenologi dan Parameterisasi Kultivar Individual.” Berdasarkan temuan dari literatur, kisaran suhu optimal untuk rami lebih rendah daripada kedelai untuk kedua proses pengembangan untuk fase vegetatif dan reproduktif (Tabel 5 ). Hal ini memungkinkan perkembangan simpul yang lebih cepat, kemajuan melalui fase fenologi, dan fotosintesis daun yang lebih tinggi pada suhu yang lebih rendah. Sensitivitas suhu pembentukan polong dan pertumbuhan biji (FNPDT, FNSDT), yang menentukan potensi hasil gabah, sedikit diturunkan berdasarkan literatur dan pembentukan biji yang diamati dalam percobaan.
| Proses Vegetatif atau Reproduksi | Tb (°C) | Puncak 1 (°C) | Suhu 2 (°C) | T maks (°C) | Referensi |
|---|---|---|---|---|---|
| Perkembangan simpul vegetatif | 3.0 | 24.0 | 29.0 | 40.0 | Van der Werf dan kawan-kawan ( 1995 ) |
| [7.0] | [28.0] | [35.0] | [45.0] | ||
| Perkembangan reproduksi awal | 4.0 | 20.0 | 30.0 | 40.0 | Amaducci dan kawan-kawan ( 2008 ) |
| [6.0] | [26.0] | [30.0] | [45.0] | ||
| Perkembangan reproduksi akhir | 4.0 | 22.0 | 31.0 | 40.0 | Amaducci dan kawan-kawan ( 2008 ) |
| [−15,0] | [26.0] | [34.0] | [45.0] | ||
| Set Pod (FNPDT) a | 10.0 | 20.0 | 26.0 | 40.0 | Dikalibrasi dari pembentukan benih yang diamati. |
| [13.0] | [21.0] | [26.5] | [40.0] | ||
| Pertumbuhan Benih (FNSDT) a | 5.0 | 18.0 | 24.0 | 42.6 | Geneve dan kawan-kawan ( 2022 ) |
| [6.0] | [21.0] | [23.5] | [41.0] | ||
| Fotosintesis daun pada CO2 tinggi (XLMAXT) | 7.0 | 35.0 | 40.0 | 50.0 | Chandra dkk. ( 2008, 2011 ), Tang dkk. ( 2017 ) |
| [7.5] | [40.0] | [44.0] | [48.0] | ||
| Perluasan luas daun (XSLATM) b | 0.0 | 10.0 | 20.0 | – | Dikalibrasi dari pengamatan perluasan luas daun pada awal musim. |
| [0.0] | [12.0] | [22.0] | [–] |
Catatan : Keempat kolom menunjukkan suhu dasar (Tb), suhu optimum pertama (Topt1), suhu optimum kedua (Topt2), dan suhu maksimum (T max ). a Empat suhu kardinal untuk pembentukan polong dan pertumbuhan biji diubah menjadi nilai proses melalui fungsi parabola empat sisi menggunakan suhu udara per jam di DSSAT. Semua fungsi lainnya menghitung laju dengan pencarian linear dengan interpolasi antara suhu kardinal menggunakan suhu udara per jam. b Ekspansi relatif 0,25 pada 0°C, 0,35 [0,25] pada 10 [12]°C dan 1,0 pada 20 [22]°C. “–” tidak ada suhu langit-langit yang digunakan.
3.1.4 Fotosintesis daun
Ganja memiliki potensi produksi biomassa tinggi dengan penutup tanah awal dan intersepsi radiasi tinggi (Struik et al., 2000 ; Van der Werf et al., 1996 ). Kultivar ganja dioecious yang ditanam dalam percobaan studi ini dicirikan oleh penuaan tanaman jantan setelah berbunga, yang sebagian dikompensasi oleh pertumbuhan berkelanjutan pada batang dan, pada tingkat yang lebih rendah, pada biomassa daun dari tanaman betina yang tersisa sepanjang periode reproduksi. Untuk tujuan pemodelan dan pengumpulan data, tanaman jantan yang menua dianggap tidak lagi menjadi bagian dari biomassa, meskipun pada tahap awal setelah berbunga, batang jantan yang sekarat akan tetap berdiri di ladang untuk beberapa waktu hingga tumbang. Untuk produktivitas biomassa umum, beberapa parameter fotosintesis dan respons N dimodifikasi untuk menyesuaikan total produksi bahan kering ganja agar sesuai dengan pola pertumbuhan biomassa yang diamati (Tabel 6 ). Luas daun spesifik acuan (SLAREF) dan berat daun spesifik untuk fotosintesis maksimum (SLWREF) ditempatkan di titik tengah antara kedelai dan kapas untuk mencapai fotosintesis dan produksi biomassa yang dibutuhkan selama musim. Laju asimilasi yang disimulasikan pada siang hari adalah 1,0–1,5 mg CO2 m − 2 s −1 pada tingkat daun, yang berada dalam kisaran wajar dari nilai yang diamati oleh Tang et al. ( 2017 ) hingga 1,4 mg CO2 m − 2 s −1 . Parameter untuk mobilisasi N umum dari jaringan vegetatif (NMOBMX) dan laju relatif selama fase vegetatif (NVSMOB) ditingkatkan untuk mencerminkan penjarangan diri dan penuaan tanaman jantan yang diamati sepanjang musim tanam. Parameter untuk penuaan daun yang lebih rendah karena naungan diri yang berlebihan (ICMP, TCMP) ditetapkan pada titik tengah antara kedelai dan kapas untuk mendukung penuaan daun umum karena penjarangan diri dan penuaan tanaman jantan. Penuaan daun pasca-pembungaan (SENRTE2) juga ditetapkan antara kedelai dan kapas untuk mencapai penurunan tajam yang diamati dalam biomassa daun pada akhir musim ketika semua atau sebagian besar gabah matang dan daun, terutama dari tanaman jantan, mengalami penuaan. Sebagai perbandingan, kedelai adalah tanaman yang mengalami penuaan diri sepenuhnya (termasuk gugurnya tangkai daun), sedangkan kapas tidak mengalami penuaan diri setelah matang. Penuaan yang diamati dalam uji lapangan terutama didorong oleh penuaan tanaman jantan setelah berbunga dan, sampai batas tertentu, penuaan daun umum selama pematangan gabah. Tidak ada kultivar rami monoecious yang menjadi bagian dari pengembangan model awal ini, tetapi memasukkannya kemungkinan akan memerlukan peninjauan ulang parameter penuaan karena tidak adanya penuaan tanaman jantan yang lengkap.
| Kelompok proses | Koefisien genetik | Singkatan dan satuan | Rami | Kedelai | Kapas |
|---|---|---|---|---|---|
| Fotosintesis | Berat daun spesifik untuk pengukuran LFMAX | SLWREF, gm −2 | 0,0046 pukul 0,0046 | 0,0036 hari | 0,0050 |
| Kemiringan berat daun spesifik (dengan kedalaman tajuk) sebagai fungsi indeks luas daun | SLWSLO, gm -2 | 0,0004 | 0,0004 | 0,0005 | |
| Kemiringan konsentrasi N daun (dengan kedalaman tajuk) sebagai fungsi indeks luas daun | LERENG, m 2 m −2 | 0.30 | 0.30 | 0,25 | |
| Nilai N daun yang di atas nilai tersebut fotosintesis daun mencapai maksimum | LNREF, % | 4.9 | 4.9 | 4.87 | |
| Pertumbuhan daun | Luas daun spesifik (SLA) saat muncul | FINREF, cm 2 g −1 | 220 | 180 | 150 |
| SLA untuk daun baru selama puncak pertumbuhan vegetatif | SLAREF, cm 2 g −1 | 240 | 350 | 170 | |
| Ukuran referensi daun simpul atas reguler | SIZREF, cm 2 daun −1 | 220 | 171.4 | 280 | |
| Tahap vegetatif yang melampaui perluasan luas daun yang dibatasi oleh penyerapan air tidak lagi dapat membatasi fotosintesis atau pertumbuhan luas daun | VSSINK | 3.0 | 5.0 | 5.0 | |
| SLA maksimum untuk daun baru bila ditanam dalam radiasi rendah tetapi air dan suhu optimal | SLAMAX, cm2g − 1 | 350 | 950 | 540 | |
| SLA minimum untuk daun baru bila tumbuh di bawah radiasi yang sangat tinggi dan air serta suhu yang optimal | SLAMIN, cm2g − 1 | 200 | 250 | 110 | |
| Penuaan daun | Faktor yang menentukan penuaan daun adalah perkalian kehilangan protein daun. | SENRTE, g(daun) g(kehilangan protein) −1 | 1.0 | 0.8 | 1.0 |
| Faktor yang menyebabkan penurunan berat daun akibat penuaan setiap hari setelah kematangan fisiologis | SENRTE2, pecahan per hari kalender | 0,04 | 0.20 | 0,01 | |
| Titik kompensasi cahaya untuk penuaan daun bagian bawah akibat naungan berlebihan oleh tajuk tanaman | ICMP, mol m −2 hari −1 | 0.4 | 0.8 | 0,01 | |
| Konstanta waktu untuk penuaan daun bagian bawah karena naungan berlebihan dari tajuk tanaman | TCMP, hari termal | 30.0 | 10.0 | 50.0 | |
| Laju mobilisasi N dari jaringan vegetatif | NMOBMX, pecahan per hari | 0,15 | 0,09 | 0,08 | |
| Laju relatif penambangan N selama fase vegetatif | NVSMOB, pecahan dari NMOBMX | 0.40 | 0,35 | 0.38 |
3.1.5 Pertumbuhan akar, penyerapan nitrogen, dan parameter stres
Akar rami dicirikan oleh pertumbuhan yang cepat dan dalam, dengan potensi penyerapan tinggi yang membuat tanaman ini cocok untuk fitoremediasi (Linger et al., 2002 ; Small, 2015 ). Untuk CROPGRO-Hemp, beberapa parameter yang menentukan pertumbuhan akar dan serapan N ditingkatkan, lebih dekat ke, atau identik dengan, parameter model CROPGRO-Cotton untuk meningkatkan ketersediaan N dan air dan untuk mencocokkan pola pertumbuhan rami yang diamati. Kedalaman akar pada hari munculnya tanaman (RTDEPI, cm) ditingkatkan dari 20 menjadi 25; panjang akar per satuan berat akar (RFAC1, cm g −1 ) ditingkatkan dari 7500 menjadi 11.000; dan serapan NO 3 dan NH 4 per panjang akar (RTNO3 dan RTNH4, mg N cm −1 hari −1 ) ditingkatkan dari 0,006 menjadi 0,010. Kesuburan tanah dan nutrisi tanaman merupakan prioritas penelitian yang sedang berlangsung untuk rami industri, karena stres abiotik dan defisit N membatasi hasil dengan mengurangi metabolisme sel, diferensiasi, dan pertumbuhan tanaman secara keseluruhan (Landi et al., 2019 ). Dalam CROPGRO, faktor stres N (NSTRES, 0–1 di mana nilai di bawah 1 berarti bahwa
) dihitung sebagai fungsi dari pasokan N dibagi dengan permintaan N pada rata-rata berjalan 2 hari. Permintaan N didorong oleh fotosintesis harian dan dapat bervariasi secara signifikan dengan variasi cuaca harian. Pasokan N bergantung pada konsentrasi N tanah dan kepadatan panjang akar di berbagai horizon tanah. Khusus untuk akar, NSTRES dapat menyebabkan peningkatan partisi ke akar yang dimodulasi oleh faktor ambang NSTFAC (0,6 untuk rami dan kapas, 0,7 untuk kedelai) yang memungkinkan peningkatan partisi ke akar. Lebih jauh, NSTRES mengurangi luas daun spesifik melalui penebalan daun (NSLA), serta tinggi dan lebar tajuk melalui pemendekan ruas (NHGT). NSLA tetap pada nilai default 1,0, tetapi NHGT ditingkatkan menjadi 5,0 untuk simulasi akurat tinggi tajuk bawah untuk eksperimen N nol. Fotosintesis daun tidak segera dikurangi oleh faktor NSTRES tetapi dikendalikan oleh konsentrasi N jaringan daun. Bahasa Indonesia: Selain defisit N, aplikasi N berlebih ditemukan mengurangi kualitas serat dan menghambat fotosintesis dan pertumbuhan melalui pengeritingan daun (Anderson et al., 2021 ), tetapi mekanisme untuk toksisitas kelebihan pasokan N saat ini tidak termasuk dalam model CROPGRO. Koefisien yang terkait dengan perluasan daun yang berkurang karena stres air (TURSLA), berkurangnya luas daun spesifik di bawah PAR tinggi (SLAPAR), gugurnya daun karena stres air (SENDAY) atau suhu rendah (FREEZ1, FREEZ2) serta penyimpanan dan mobilisasi karbohidrat (CMOBMX, CADSTF, dan CADPRI1), tetap seperti pada kedelai. Parameter terkait stres ini bukan fokus pengembangan model awal ini karena datanya terbatas untuk kalibrasi. Eksperimen di masa mendatang mungkin memberikan pemahaman yang lebih mendalam tentang respons stres N, air, dan abiotik rami dan akan memungkinkan kalibrasi yang lebih tepat dari parameter ini untuk memodelkan produksi di bawah skenario stres.
3.1.6 Fenologi dan parameterisasi kultivar individu
Rami adalah tanaman hari pendek fakultatif, dengan fotoperiode dan suhu mendorong perkembangan fenologi. Kultivar dioecious yang digunakan dalam penelitian ini memiliki tanaman jantan yang menghasilkan serbuk sari dan tanaman betina yang menghasilkan bunga yang berkembang menjadi biji. Kemajuan menuju pembungaan lebih lambat, dan tahap vegetatif dipertahankan jika panjang hari di atas ambang batas fotoperiode kritis (Moher et al., 2021 ). M. Zhang et al. ( 2021 ) menemukan efek fotoperiode yang kuat, yang berarti bahwa perbedaan panjang hari tambahan sedikitnya 15 menit setelah dimulainya fotoperiode kritis dapat menunda respons pembungaan hingga 5–13 hari tambahan. Lebih jauh lagi, intensitas cahaya yang relatif rendah, misalnya, senja sipil sebelum matahari terbit atau setelah matahari terbenam, ditemukan cukup untuk menjadi aktif secara biologis dan mengatur respons pembungaan (M. Zhang et al., 2021 ). Persyaratan fotoperiodik yang berbeda di antara kultivar kemungkinan didorong oleh kondisi fotoperiodik lokal di garis lintang asal sebagai adaptasi terhadap panjang hari dan musim yang terjadi secara alami (De Meijer & Keizer, 1994 ; Ranalli, 1999 ). Transisi ke pembungaan memperlambat pertumbuhan tinggi dan biomassa, mengurangi populasi tanaman kultivar dioecious melalui penuaan tanaman jantan, dan dikaitkan dengan puncak kualitas serat dan waktu panen yang direkomendasikan untuk penggunaan serat. Pematangan dan pengerasan batang serat yang terus menerus setelah pembungaan membuatnya kurang cocok untuk pemrosesan industri (Salentijn et al., 2019 ) meskipun penelitian lebih lanjut dalam kultivar yang mempertahankan kualitas serat selama pembungaan atau pematangan benih sedang berlangsung (Petit et al., 2020 ). Dalam pemodelan berbasis proses, prediksi permulaan dan durasi fase fenologi, oleh karena itu, penting untuk prediksi dinamika hasil dan biomassa sepanjang musim.
Dalam CSM dan CROPGRO, fase fenologi dipisahkan menjadi beberapa fase vegetatif dan reproduktif dari kemunculan hingga kematangan fisiologis, dengan persyaratan PD khusus untuk maju dari satu fase ke fase berikutnya. PD sama dengan hari kalender pada suhu dan panjang hari yang optimal. Penting untuk mempertimbangkan bahwa panjang hari dalam CROPGRO dan CSM didefinisikan sebagai waktu dari matahari terbit hingga terbenam ketika sinar matahari dapat langsung menyinari tajuk. Matahari terbit adalah ketika cabang atas matahari tampak bersinggungan dengan cakrawala, sedangkan matahari terbenam adalah turunnya matahari di bawah cakrawala. Waktu senja atau persyaratan intensitas cahaya khusus spesies untuk memicu respons terang/gelap tidak dipertimbangkan untuk perhitungan panjang hari dalam CSM dan CROPGRO. Untuk tanaman hari pendek, fotoperiode memodulasi kemajuan fenologi dengan memperlambat perkembangan jika panjang hari saat ini berada di atas CSDL khusus kultivar. PP-SEN mendefinisikan kemiringan respons, dengan nilai yang lebih tinggi mengarah pada penundaan yang lebih lama dalam perkembangan. Pertumbuhan dalam kondisi hari panjang akan menghasilkan periode vegetatif yang berkelanjutan, yang sering kali diinginkan untuk mencapai pertumbuhan biomassa yang cukup sebelum dipanen untuk keperluan serat di sekitar waktu pembungaan. Untuk pembentukan biji rami, CROPGRO menggunakan istilah polong, kulit, dan biji yang berasal dari kedelai. Untuk CROPGRO-Hemp, istilah “biji” mengacu pada biji rami yang sebenarnya dan istilah “kulit” mengacu pada bagian tanaman yang masih berbunga (cola), seperti daun kecil (bracts) dan struktur pendukung. “Polong” adalah gabungan dari biji dan kulit.
Kultivar yang berbeda diparameterisasi melalui seperangkat parameter kultivar dan ekotipe. Terkait dengan konsep ekotipe ekologi (Quinn, 1978 ), parameter ekotipe umumnya berlaku untuk seluruh populasi dari area tertentu dengan karakteristik yang berbeda. Satu set parameter ekotipe dapat berlaku untuk beberapa kultivar, meskipun ini tidak terjadi dalam penelitian kami. Kami menemukan perbedaan yang signifikan dalam perkembangan simpul, tinggi tajuk, fenologi, dan pembentukan biji antara dua kultivar awal, yang menghasilkan set parameter ekotipe yang sedikit berbeda untuk setiap kultivar. Beberapa parameter kultivar (Tabel 7 ) dan ekotipe (Tabel 8 ) disesuaikan untuk mewakili perilaku pembungaan dan pembentukan biji yang berbeda dari IH Williams dan NWG 2730, dengan yang terakhir umumnya berbunga lebih awal dan menunjukkan perkembangan yang lebih cepat ke pembentukan biji dan penuaan daun di akhir musim. Dimulai dengan penaburan, tanaman rami muncul setelah sejumlah hari minimum yang ditetapkan dari penanaman hingga kemunculan (PL-EM), yang dikalibrasi pada 4,5 PD berdasarkan kemunculan yang diamati pertama kali. Tanaman kemudian menjalani rentang waktu yang singkat dari kemunculan hingga daun sejati pertama (EM-V1) di mana mereka masih muda dan induksi bunga tidak memungkinkan. Induksi bunga hanyalah kompetensi meristem epikal untuk mengembangkan bunga, tetapi bukan penampilan bunga yang sebenarnya terlihat. Itu tidak diukur tetapi hanya diperkirakan dan dapat didefinisikan ulang berdasarkan wawasan dari pengamatan jaringan meristem. Waktu dari daun sejati pertama hingga akhir fase muda (V1-JU) ditetapkan pada 0, mengakhiri fase muda setelah daun sejati pertama dan memulai kemungkinan induksi bunga. Jumlah minimum 1,0 hari ditetapkan untuk jumlah PD untuk induksi bunga di bawah suhu dan panjang hari yang optimal (JU-R0) berdasarkan wawasan bahwa rami dapat beralih dengan cepat ke pembungaan jika, misalnya, kondisi cahaya berubah dalam lingkungan yang terkendali.
| Koefisien genetik | Rami | Rami | Kedelai | Kapas |
|---|---|---|---|---|
| Nama kultivar | IH Williams | Kelompok Kerja Nasional 2730 | Bragg | Deltapin 77 |
| Panjang hari kritis (CSDL) | pukul 13.45 | pukul 13.55 | 12.33 | pukul 23.00 |
| Respons relatif perkembangan terhadap fotoperiode (PPSEN) | 0,90 | 0.32 | 0.32 | 0,01 |
| Waktu (PD) dari kemunculan hingga pembungaan (EM-FL) | 20.0 | 16.5 | 19.5 | 34.0 |
| Waktu (PD) dari pembungaan hingga cangkang pertama (FL-SH) | 9.0 | 6.0 | 10.0 | 8.0 |
| Waktu (PD) dari pembungaan hingga benih pertama (FL-SD) | 15.0 | 10.0 | 15.2 | 15.0 |
| Waktu (PD) dari benih pertama hingga kematangan fisiologis (SD-PM) | 53.0 | 30.0 | 37.6 | 49.0 |
| Waktu (PD) dari pembungaan hingga pertumbuhan daun akhir (FL-LF) | 30.0 | 25.0 | 19 | 75 |
| Fotosintesis daun maksimum pada suhu tinggi, CO 2, dan cahaya (LFMAX, mg CO 2 m − 2 s − 1 ) | 1.30 | 1.30 | 1.00 | 1.10 |
| Luas daun spesifik (SLAVR, cm 2 g − 1 ) | 240 | 230 | 355 | 170 |
| Ukuran maksimum daun penuh (SIZLF, cm 2 ) | 220 | 220 | 170 | 280 |
| Fraksi maksimum pertumbuhan harian dibagi ke dalam biji + cangkang (XFRT) | 0.40 | 0,70 | 1.00 | 0,80 |
| Berat maksimum per biji (WTPSD, g) | 0,02 | 0,02 | 0.17 | 0.18 |
| Durasi pengisian benih per kelompok polong (SFDUR, PD) | 20 | 20 | 24 | 35 |
| Rata-rata jumlah benih per polong (SDPDV, jumlah per polong) | 1 | 1 | 2 | 27 |
| Waktu (PD) yang dibutuhkan untuk mencapai beban pod akhir (PODUR) | 30 | 25 | 10 | 8 |
| Rasio maksimum (biji (biji + kulit) −1 ) pada saat matang a (THRSH) | 80 | 80 | 78 | 70 |
| Fraksi protein dalam biji (SDPRO, g(protein) g(biji) − 1 ) | 0.256 | 0.256 | 0.4 | 0.153 |
| Fraksi minyak dalam biji (SDLIP, g(minyak) g(biji) − 1 ) | 0.319 | 0.319 | 0.2 | 0.12 |
Singkatan: PD, hari fototermal.
a Menyebabkan benih berhenti tumbuh karena berat keringnya meningkat hingga cangkang terisi dalam satu kelompok.
| Koefisien genetik | Rami | Rami | Kedelai | Kapas |
|---|---|---|---|---|
| Ekotipe | HM0001 | HM0002 | Nomor SB0701 | Deltapin 77 |
| Keterangan | IH Williams | Kelompok Kerja Nasional 2730 | Kedelai | DP 77 |
| Jumlah hari minimum (PD) dari penanaman hingga kemunculan (PL-EM) | 4.5 | 4.5 | 2.2 | 4 |
| Jumlah hari minimum (PD) dari kemunculan hingga daun sejati pertama (EM-V1) | 3 | 3 | 6 | 4 |
| Jumlah hari minimum (PD) dari daun sejati pertama hingga akhir fase juvenil (V1-JU) | angka 0 | angka 0 | angka 0 | angka 0 |
| Jumlah hari minimum (PD) untuk induksi bunga (JU-R0) | 1 | 1 | 5 | angka 0 |
| Proporsi waktu antara benih pertama dan kematangan fisiologis dimana benih terakhir dapat terbentuk (PM09) | 0,80 | 0,80 | 0,35 | 0,90 |
| Waktu (PD) yang dibutuhkan untuk pertumbuhan cangkang individu (LNGSH) | 5 | 5 | 10 | 10 |
| Waktu (hari) antara kematangan fisiologis dan panen (R7–R8) | 25 | 20 | 12 | 10 |
| Waktu (PD) dari bunga pertama hingga daun terakhir pada batang utama (FL-VS) | 53 | 34 | 9 | 75 |
| Tingkat kemunculan nodus pada batang utama (TRIFL, nodus PD − 1 ) | 0.28 | 0.31 | 0.32 | 0.33 |
| Tinggi ekotipe ini terhadap tinggi standar (RHGHT) | 1.20 | 0,80 | 1.00 | 0,85 |
Catatan : Koefisien yang terkait dengan kelompok kematangan (MG) dan adaptasi suhu (TM) tidak berlaku untuk rami dan ditetapkan ke 1.
Singkatan: PD, hari fototermal.
Bahasa Indonesia: Untuk pembungaan yang terlihat, metode yang berbeda tersedia untuk mengklasifikasikan fenologi pembungaan rami, dan kalibrasi model ditargetkan pada pengamatan “bunga terminal pertama” (Brym et al., 2023 ). Dalam percobaan yang disertakan untuk pengembangan model, 50% pembungaan umumnya terjadi 2–3 minggu setelah tanda-tanda pertama yang terlihat dari pembungaan soliter/aksiler, yaitu 1–2 minggu setelah pembungaan terminal pertama. Bunga terakhir terlihat 2–3 minggu setelah 50% pembungaan, yang mengarah ke total periode pembungaan 4–6 minggu. Dalam CROPGRO-Hemp, setelah induksi, pembungaan dimulai setelah rentang waktu minimum dari kemunculan hingga pembungaan (EM-FL) terpenuhi. EM-FL ditetapkan pada 20,0 PD untuk IH Williams dan diturunkan menjadi 16,5 PD untuk NWG 2730 untuk membuat ambang batas yang lebih rendah untuk tanggal pembungaan paling awal yang diamati dalam kondisi hari pendek berkelanjutan dan suhu optimal. Efek fotoperiode dapat memperpanjang jangka waktu minimum ini berdasarkan CSDL spesifik kultivar dan kemiringan respons (PPSEN), yang ditetapkan untuk menyesuaikan perbedaan tanggal pembungaan antara lokasi dan tanggal penanaman yang berbeda.
Dengan tidak adanya data eksperimen terperinci, baik CSDL maupun PPSEN dikalibrasi secara empiris. Eksperimen di lokasi WFREC yang lebih utara umumnya memiliki waktu yang sedikit lebih lama untuk berbunga dibandingkan dengan PSREU. CSDL ditetapkan berdasarkan perbedaan panjang hari pada bulan Mei antara PSREU dan lokasi WFREC sehingga eksperimen utara di WFREC mengalami kondisi hari panjang setelah penanaman dan mereplikasi waktu yang diperpanjang dari penanaman hingga pembungaan yang diamati di WFREC, terutama untuk penanaman akhir Mei di musim 2021. Penundaan tambahan dalam pembungaan saat berpindah dari lokasi PSREU ke lokasi WFREC yang lebih utara lebih besar untuk IH Williams dibandingkan dengan NWG 2730. PPSEN disesuaikan berdasarkan besarnya penundaan pembungaan ini dan ditetapkan secara signifikan lebih tinggi untuk kultivar IH Williams. Selain itu, CSDL dan PPSEN disetel dengan baik menggunakan pengamatan fenologi tambahan dari garis lintang yang lebih utara dengan rentang waktu yang jauh lebih lama antara penanaman dan pembungaan. CSDL yang diperkirakan sebesar 13,45 jam untuk IH Williams dan 13,55 jam untuk NWG 2730 berada di bawah CSDL yang diukur untuk kultivar rami lain dalam penelitian sebelumnya (M. Zhang et al., 2021 ). Hal ini dapat disebabkan oleh perbedaan definisi panjang hari dengan atau tanpa senja atau cahaya intensitas sangat rendah. Model ini dikalibrasi untuk panjang hari dari matahari terbit hingga terbenam, tanpa mempertimbangkan senja atau cahaya intensitas rendah lainnya. Atau, CSDL untuk IH Williams dan NWG 2730 memang bisa jadi jauh lebih rendah daripada yang diukur untuk kultivar lain, yang sebagian menjelaskan mengapa IH Williams dan NWG 2730 tampak lebih cocok dan produktif di garis lintang selatan seperti Florida karena periode vegetatif yang diperpanjang. Validasi respons fotoperiode yang diidentifikasi secara empiris dengan eksperimen spesifik direkomendasikan.
Setelah berbunga, cangkang dan biji pertama terbentuk dengan penundaan 9,0/15,0 PD untuk IH Williams dan 6,0/10,0 PD untuk NWG 2730 (FL-SH/FL-SD). Beberapa parameter biji dan pematangan disesuaikan untuk memperpanjang periode pembungaan dan perkembangan biji dan mencocokkan pola pembungaan tak tentu yang diamati, di mana bunga dan biji dengan kematangan berbeda dapat hadir pada tanaman yang sama pada waktu yang sama (Elias et al., 2020 ). PM09 menetapkan proporsi waktu antara biji pertama dan kematangan fisiologis di mana biji terakhir dapat terbentuk dan ditingkatkan menjadi 0,80 untuk memperpanjang dan meniru fase pembungaan tak tentu dan pembentukan biji rami. XFRT ditetapkan sebagai parameter partisi konstan antara 0 dan 1, dengan 1 menunjukkan bahwa semua asimilat dapat dialokasikan untuk pertumbuhan reproduktif jika jumlah biji maksimum tercapai. Nilainya ditetapkan sebesar 0,40 untuk IH Williams dan 0,70 untuk NWG 2730, dan benih memiliki prioritas jika benih yang terbentuk cukup untuk menyerap asimilat. Asimilat yang tersisa yang tidak digunakan untuk pertumbuhan reproduksi dialokasikan mengikuti pembagian asimilat yang bergantung pada “Tahap-V” yang dijelaskan sebelumnya ke organ vegetatif.
Koefisien kultivar PODUR mendefinisikan waktu fototermal yang diperlukan bagi tanaman untuk mencapai beban polong akhir dan ditingkatkan menjadi 30,0 PD untuk IH Williams untuk mereplikasi penambahan polong terus-menerus selama fase pembungaan tak tentu. Untuk NWG 2730, PODUR ditetapkan sedikit lebih rendah pada 25,0 karena pembentukan benih yang diamati sebelumnya. Koefisien kultivar SDPDV mendefinisikan jumlah benih per polong dan ditetapkan sebesar 1,0, menghasilkan satu benih per polong tetapi sejumlah besar polong per tanaman. Durasi pengisian benih dari benih individu (SFDUR) ditetapkan pada 20 PD untuk mereplikasi durasi pengisian gabah yang diamati selama periode sekitar 30 hari kalender. Perluasan luas daun yang berkelanjutan setelah bunga pertama (FL-LF) dikalibrasi untuk mempertahankan perluasan luas daun untuk waktu yang terbatas pasca-pembungaan hingga peningkatan tajam dalam penuaan daun terjadi pada akhir pembungaan dan awal pematangan benih.
Total waktu dari benih pertama hingga kematangan fisiologis (SD-PM) ditetapkan pada 53 PD untuk IH Williams dan 30 untuk NWG 2730 untuk mencapai kematangan fisiologis benih pertama dan peningkatan kuat berikutnya dalam penuaan daun menjelang akhir musim tanam. PD antara kematangan fisiologis dan panen (R7–R8) ditetapkan pada 25 untuk IH Williams dan 20 untuk NWG 2730 untuk memberikan benih yang terbentuk akhir waktu untuk meningkatkan berat kering dan matang. Fraksi minyak dan protein dalam benih (SDPRO, SDLIP) diasumsikan sama untuk IH Williams dan NWG 2730 tetapi dapat disesuaikan untuk mencerminkan perbedaan kultivar yang diketahui (Vonapartis et al., 2015 ). Dalam kerangka CROPGRO umum, durasi berbagai tahap fenologi dapat dipersingkat atau diperpanjang lebih lanjut oleh stres air, N, atau fosfor, tetapi fitur-fitur ini dinonaktifkan untuk Hemp karena kurangnya data yang tersedia untuk kalibrasi. Perubahan yang dibahas sebelumnya menyelesaikan kalibrasi parameter model CROPGRO-Hemp untuk satu serat dan satu kultivar benih. Gambaran umum parameter model lengkap, termasuk parameter yang tidak diubah selama proses pengembangan, tersedia di Informasi Pendukung 1 .
3.2 Tinjauan kinerja model dan hasil simulasi
3.2.1 Fenologi
Simulasi tanggal berbunga dibandingkan dengan pengamatan untuk “bunga terminal pertama” dan mencapai ini secara subjektif dengan baik di semua simulasi. Deviasi maksimum adalah 5 hari terlambat hingga 3 hari lebih awal di semua eksperimen untuk kedua kultivar, dua musim, dan dua lokasi di Florida (Tabel 9 ). Mengingat rentang lintang yang terbatas di kedua lokasi, pengamatan fenologi tambahan untuk dua kultivar yang sama untuk lokasi yang lebih jauh di utara di Illinois, Michigan, dan Kansas, dengan waktu yang jauh lebih lama untuk berbunga, disertakan untuk evaluasi awal. Pengamatan diperoleh dari laporan eksperimen dan Midwestern Hemp Database (Alberti, 2021 ; University of Illinois, 2024 ). Di lokasi di luar Florida, tanggal berbunga yang disimulasikan hingga 4 hari lebih awal di Illinois dan 4 hari lebih lambat di Michigan untuk kultivar NWG 2730 dan hingga 3 hari lebih awal untuk kultivar IH Williams di Illinois. Ini masih dapat diterima mengingat spesifikasi tanggal berbunga yang tidak pasti dan mempertimbangkan bahwa perbedaan antara 50% dan pembungaan terminal dapat sekitar 10 hari.
| Berbunga | |||||||
|---|---|---|---|---|---|---|---|
| Kultivar | Lokasi | Musim | Penanaman | 50% | Terminal | Disimulasikan | Perbedaan b |
| IH Williams | PSREU | Tahun 2021 | 6 Mei | 16 Juni | 6 Juni | 6 Juni | angka 0 |
| IH Williams | PSREU | Tahun 2022 | 5 Mei | 12 Juni | 2 Juni | 3 Juni | +1 |
| IH Williams | Bahasa Inggris WFREC | Tahun 2021 | 26 Mei | 15 Juli | 5 Juli | 6 Juli | +1 |
| IH Williams | Bahasa Inggris WFREC | Tahun 2022 | 13 Mei | 28 Juni | 18 Juni | 19 Juni | +1 |
| Kelompok Kerja Nasional 2730 | PSREU | Tahun 2021 | 6 Mei | 8 Juni | 28 Mei | 29 Mei | +1 |
| Kelompok Kerja Nasional 2730 | PSREU | Tahun 2022 | 5 Mei | 4 Juni | 24 Mei | 26 Mei | +2 |
| Kelompok Kerja Nasional 2730 | Bahasa Inggris WFREC | Tahun 2021 | 26 Mei | 5 Juli | 25 Juni | 20 Juni | -5 |
| Kelompok Kerja Nasional 2730 | Bahasa Inggris WFREC | Tahun 2022 | 13 Mei | 14 Juni | 4 Juni | 5 Juni | +1 |
| IH Williams | Kota Urbana, Illinois | Tahun 2023 | 18 Juni | 6 Agustus | 27 Juli | 29 Juli | +2 |
| IH Williams | Manhattan, KS sebuah | Tahun 2022 | Tanggal 20 april | 19 Agustus | 9 Agustus | 11 Agustus | +2 |
| IH Williams | Hyde, Michigan, Amerika Serikat | Tahun 2023 | 9 Juni | 11 Sept. | 1 Sept | 1 Sept | angka 0 |
| Kelompok Kerja Nasional 2730 | Kota Urbana, Illinois | Tahun 2021 | 3 Juni | – | 16 Juli | 13 Juli | -3 |
| Kelompok Kerja Nasional 2730 | Kota Urbana, Illinois | Tahun 2022 | 24 Mei | – | 11 Juli | 7 Juli | -4 |
| Kelompok Kerja Nasional 2730 | Manhattan, Kansas | Tahun 2020 | 8 Juni | – | 11 Juli | 13 Juli | +2 |
| Kelompok Kerja Nasional 2730 | Manhattan, Kansas | Tahun 2021 | 4 Juni | – | 12 Juli | 9 Juli | -3 |
| Kelompok Kerja Nasional 2730 | Hyde, Michigan, Amerika Serikat | Tahun 2021 | 27 Mei | – | 26 Juli | 30 Juli | +4 |
Catatan : Situs PSREU adalah Plant Science Research and Education Unit di Citra, FL. Situs WFREC adalah West Florida Research and Education Center di Jay, FL. Referensi untuk data pembungaan dari luar Florida dalam urutan baris tabel: Alberti ( 2021 ), Dedecker et al. ( 2021 ), GHA ( 2022 ), Lee et al. ( 2022 ), Roozeboom dan Griffin ( 2021, 2023a ), dan University of Illinois ( 2024 ). Metodologi yang tepat untuk penentuan pembungaan tidak diketahui untuk kultivar IH Williams di lokasi di luar Florida tetapi diasumsikan pada 50% pembungaan. Pengamatan pembungaan untuk NWG 2730 di lokasi di luar Florida digambarkan sebagai "bunga pertama yang terlihat" atau "kuncup pertama," yang kami asumsikan sebanding dengan "pembungaan terminal." a Lokasi pasti di Kansas untuk uji coba IH Williams tidak diketahui karena hanya negara bagiannya saja yang disebutkan. Manhattan, Kansas diasumsikan karena adanya penelitian rami lain di lokasi ini oleh Roozeboom dan Griffin ( 2023b ). b Perbedaan dihitung antara simulasi pembungaan dan pembungaan terminal yang diamati.
Bahasa Indonesia: Kami mengakui bahwa simulasi fenologi rami dengan data cuaca yang diperoleh dari satelit dan lokasi uji coba yang tidak diketahui secara pasti memiliki ketidakpastian, tetapi kami menganggap perbandingan tersebut berharga untuk menggarisbawahi bahwa simulasi fenologi tampaknya secara umum masuk akal di luar dua lokasi di Florida. Lebih jauh lagi, akurasi umum pengamatan fenologi sering dibatasi oleh frekuensi kunjungan lapangan dua mingguan hingga mingguan dan kesulitan dalam menentukan tahap pembungaan rami secara akurat. Sebagai perbandingan, tinjauan ekstensif di antara 27 kelompok pemodelan untuk fenologi gandum, tanaman yang dipelajari lebih mendalam dengan sejarah panjang pengembangan dan peningkatan model tanaman, menyimpulkan kesalahan absolut rata-rata sekitar 3 hari untuk simulasi fenologi gandum di Prancis (Wallach et al., 2021 ). Tahap reproduksi rami yang didefinisikan dengan cermat dan studi fenologi lingkungan yang terkendali dapat memberikan kumpulan data yang lebih baik untuk pengembangan dan evaluasi model di masa mendatang, seperti yang dicatat dalam penelitian sebelumnya tentang pemodelan fenologi rami oleh Amaducci et al. ( 2008, 2012 ).
Biomassa dan berat gabah selalu disimulasikan dan diukur pada kadar air 0%. Simulasi keseluruhan biomassa di atas tanah (atas) dan batang, berat gabah, serta tinggi tajuk menunjukkan kesesuaian yang baik antara nilai simulasi dan teramati di semua simulasi, ditunjukkan oleh nilai d-statistik yang berkisar dari 0,74 hingga 0,92 (Tabel 10 ). RMSE masuk akal jika diletakkan dalam perspektif nilai absolut observasi dengan maksimum 10.000 kg ha −1 untuk biomassa atas, 5000 kg ha −1 untuk biomassa batang, 1000 kg ha −1 untuk berat gabah, dan 2,50 m untuk tinggi tajuk. Perbandingan hasil simulasi menurut kelompok berdasarkan musim, kultivar, dan perlakuan N tidak menunjukkan perbedaan besar antara kelompok, yang menunjukkan bahwa kinerja simulasi stabil di seluruh perbedaan iklim, lokasi, fisiologis, dan manajemen yang dipertimbangkan. Hasil statistik untuk simulasi individual (Tabel 11 ) mengungkap lebih banyak variabilitas antara simulasi individual, dengan khususnya simulasi 6 ( d = 0,64, RMSE = 220 kg ha −1 ) untuk percobaan dengan kultivar NWG 2730 di lokasi PSREU pada tahun 2022 dan perlakuan 0 kg N ha −1 dan simulasi 15 ( d = 0,68, RMSE = 1010 kg ha −1 ) untuk percobaan dengan kultivar NWG 2730 di lokasi WFREC pada tahun 2022 dan perlakuan 280 kg N ha −1 , yang tidak sesuai dengan biomassa pucuk yang diamati sebaik pada simulasi lainnya.
| Biomassa teratas | Biomassa batang | Tinggi kanopi | Berat biji-bijian | |||||
|---|---|---|---|---|---|---|---|---|
| Kelompok Simulasi | d-stat | Nilai RMS (kg ha −1 ) | d-stat | Nilai RMS (kg ha −1 ) | d-stat | Nilai RMSE (m) | d-stat | Nilai RMS (kg ha −1 ) |
| Semua simulasi | 0,91 | 409 | 0,88 | 398 | 0.92 | 0.21 | 0.74 | 229 |
| Musim 2021 | 0,97 | 249 | 0,95 | 264 | 0,93 | 0.19 | 0.67 | 241 |
| Musim 2022 | 0.86 | 549 | 0.81 | 514 | 0,91 | 0.23 | 0,88 | 209 |
| Situs PSREU | 0.92 | 366 | 0,90 | 285 | 0.92 | 0.21 | 0.83 | 196 |
| Situs WFREC | 0,91 | 446 | 0.86 | 496 | 0,93 | 0.21 | 0.27 | 289 |
| Kultivar IH Williams | 0,96 | 530 | 0,90 | 639 | 0,94 | 0.22 | 0,70 | 287 |
| Kultivar NWG 2730 | 0,87 | 303 | 0,85 | 187 | 0,90 | 0.20 | 0.76 | 181 |
| Perawatan tanpa nitrogen | 0,88 | 253 | 0.86 | 214 | 0,88 | 0.24 | 0,85 | 105 |
| Perawatan nitrogen tinggi | 0,94 | 546 | 0,89 | 558 | 0,97 | 0.19 | 0.72 | 334 |
Catatan : Lokasi PSREU adalah Plant Science Research and Education Unit di Citra, FL. Lokasi WFREC adalah West Florida Research and Education Center di Jay, FL. Biomassa teratas sama dengan gabungan biomassa batang, daun, kulit, dan biji-bijian. Biomassa dan berat biji-bijian berada pada kadar air 0%.
| Biomassa teratas | Biomassa daun | Biomassa batang | Biomassa akar | Berat biji-bijian | Jumlah node | Tinggi kanopi | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Simulasi | d-stat | Nilai RMS (kg ha −1 ) | d-stat | Nilai RMS (kg ha −1 ) | [statistik d] | Nilai RMS (kg ha −1 ) | d-stat | Nilai RMS (kg ha −1 ) | d-stat | [RMSE] (kg ha -1 ) | d-stat | RMSE (angka) | d-stat | Nilai RMSE (m) |
| 1 | 0,99 | 433 | 0,99 | 150 | 0,96 | 669 | – | – | 0,60 | 339 | 0,99 | 1.30 | 0,98 | 0.22 |
| 2 | 0.92 | 181 | 0.92 | 61 | 0.86 | 145 | 0.64 | 67 | 0,85 | 51 | 0,99 | 1.00 | 0,96 | 0.11 |
| 3 | 0,98 | 293 | 0.86 | 202 | 0,99 | 116 | 0.86 | 169 | 0,95 | 144 | 0,99 | 1.33 | 0,95 | 0.23 |
| 4 | 0,95 | 248 | 0,90 | 138 | 0.86 | 330 | 0,25 | 148 | – | 259 | 0,96 | 1.93 | 0,94 | 0.22 |
| 5 | 0,96 | 926 | 0,95 | 241 | 0,96 | 532 | 0,09 | tahun 1132 | 0,79 | 397 | 0,94 | 1.64 | 1.00 | 0,07 |
| 6 | 0.64 | 220 | 0,90 | 36 | 0,65 | 131 | – | 170 | – | 66 | – | 2.10 | 0.67 | 0.43 |
| 7 | 0,98 | 264 | 0,79 | 258 | 0,98 | 75 | 0,15 | 347 | 0,96 | 113 | 0.81 | 2.11 | 0,91 | 0.23 |
| 8 | 0,99 | 128 | 0,99 | 51 | 0,98 | 122 | 0.34 | 339 | – | 1 | 0,91 | 2.53 | 0,77 | 0.38 |
| 9 | 0,99 | 439 | 0,98 | 195 | 0,97 | 520 | 0.54 | 153 | – | 440 | 0,96 | 1,99 | 0,97 | 0.22 |
| 10 | 0,94 | 81 | 0.43 | 82 | 0,99 | 15 | 0.48 | 125 | – | 145 | – | 0.30 | 0,93 | 0.10 |
| 11 | 0,98 | 189 | 0.68 | 147 | 0,89 | 265 | 0.30 | 184 | 0.27 | 568 | – | 0,65 | 0,98 | 0.10 |
| 12 | 0.86 | 724 | 0,80 | 221 | 0,78 | 701 | 0,06 | 660 | – | – | 0,95 | 1.89 | 0,97 | 0.18 |
| 13 | 0,97 | 810 | 0,90 | 356 | 0,80 | tahun 1596 | – | 744 | – | – | 0.74 | 4.29 | 0,98 | 0.26 |
| 14 | 0.84 | 188 | 0.36 | 519 | 0,90 | 57 | – | – | – | – | – | 1.70 | 0,93 | 0.23 |
| 15 | 0.68 | tahun 1010 | 0.29 | tahun 1043 | 0.56 | 694 | – | – | – | – | – | 0,75 | 0,97 | 0.17 |
Catatan : Simulasi 1–7 merujuk pada eksperimen di lokasi PSREU (Plant Science Research and Education Unit) di Citra, FL; 8–15 di lokasi WFREC (West Florida Research and Education Center) di Jay, FL; 1–3, 8–11 pada tahun 2021, 4–7 dan 12–15 pada tahun 2022; 1, 4, 5, 8, 9, 12, dan 13 untuk kultivar IH Williams, sisanya untuk kultivar NWG 2730; 1, 3, 5, 7, 9, 11, 13, dan 15 untuk perlakuan pupuk N tinggi, sisanya untuk perlakuan 0 kg N ha −1 . Rincian tambahan tersedia dalam Tabel 2. Biomassa teratas sama dengan gabungan biomassa batang, daun, dan gabah. Biomassa dan berat gabah pada kadar air 0%. “–” menunjukkan bahwa tidak mungkin menghitung d karena hanya ada nol atau satu observasi yang tersedia, atau tidak mungkin menghitung RMSE karena tidak ada observasi yang tersedia untuk jenis observasi tertentu dalam percobaan ini.
Percobaan dengan kultivar NWG 2730 di lokasi PSREU pada tahun 2022 dan perlakuan 0 kg N ha −1 yang setara dengan simulasi 6 dicirikan oleh produktivitas biomassa yang luar biasa rendah dengan mempertimbangkan perbedaan besar antara perlakuan 0 dan 56 kg N ha −1 dalam percobaan ini, dibandingkan dengan perbedaan yang lebih kecil antara perlakuan N yang sama dalam percobaan lain (N. Kaur et al., 2024 ). Biomassa di atas tanah yang disimulasikan dan diamati cocok dengan baik untuk paruh pertama musim. Oleh karena itu, kami berasumsi bahwa faktor abiotik tambahan, atau peningkatan tekanan gulma, penjarangan sendiri, atau peningkatan penuaan pasca-pembungaan karena stres pada paruh kedua musim berkontribusi terhadap penurunan biomassa yang diamati. Efek-efek ini tidak termasuk dalam model; oleh karena itu, model tersebut memprediksi secara berlebihan baik berat daun maupun batang untuk percobaan ini, yang mengarah pada prediksi kumulatif yang berlebihan dari berat pucuk.
Simulasi 15 untuk percobaan dengan kultivar NWG 2730 di lokasi WFREC pada tahun 2022 dan perlakuan 280 kg N ha −1 (Tabel 11 ) secara signifikan meremehkan biomassa di atas tanah, khususnya berat daun. Hanya dua sampel biomassa menjelang akhir musim tanam yang dikumpulkan untuk percobaan ini, yang membuat diagnosis menjadi sulit karena prediksi yang kurang pada awal musim dapat menyebar dan menghasilkan penyimpangan yang lebih besar antara simulasi dan pengamatan di akhir musim. Selain itu, ketidakpastian dalam pengamatan pembungaan dan kalibrasi fenologi berikutnya dapat mengakibatkan prediksi yang kurang serupa terhadap biomassa. Misalnya, dengan tanggal pembungaan yang disimulasikan lebih awal, periode pertumbuhan vegetatif yang disimulasikan akan lebih pendek daripada periode pertumbuhan vegetatif yang sebenarnya, yang menyebabkan timbulnya penuaan daun pasca-pembungaan lebih awal dan pertumbuhan biomassa yang berkurang secara keseluruhan.
Meskipun akurasi simulasi biomassa pucuk pada simulasi lain baik, simulasi 10–11 untuk eksperimen dengan kultivar NWG 2730 di lokasi WFREC tahun 2021 dan perlakuan 0 atau 112 kg N ha −1 serta simulasi 14–15 untuk eksperimen dengan kultivar NWG 2730 di lokasi WFREC tahun 2022 dan perlakuan 0 atau 280 kg N ha −1 (Tabel 11 ) memiliki akurasi yang bervariasi dalam simulasi masing-masing komponen biomassa daun dan batang. Penjelasan yang mungkin adalah deviasi yang disebutkan sebelumnya antara fenologi yang disimulasikan, diamati, dan sebenarnya, karena pembungaan yang disimulasikan lebih awal atau lebih lambat menggeser permulaan penuaan daun pasca-pembungaan dan menyebabkan pengurangan berat daun yang tajam. Lebih jauh, karena galat lebih jelas untuk kultivar NWG 2730, ada kemungkinan bahwa kalibrasi partisi asimilasi saat ini bias terhadap kultivar IH Williams dan tidak cukup memperhitungkan perbedaan fisiologis dalam NWG 2730. Analisis lebih lanjut dari NWG 2730 di lokasi atau musim tambahan akan memberikan gambaran yang lebih komprehensif tentang fenologi dan produksi biomassa kultivar ini untuk perbaikan model. Di semua percobaan, jumlah simpul diprediksi dengan baik ( d = 0,81–0,99) kecuali untuk simulasi 13 untuk percobaan dengan kultivar IH Williams di lokasi WFREC pada tahun 2022 dan perlakuan 280 kg N ha −1 , di mana jumlah simpul pertengahan musim yang diamati lebih tinggi daripada jumlah simpul akhir musim, kemungkinan karena kesalahan pengukuran atau pengambilan sampel yang bias.
Keakuratan untuk simulasi berat gabah sangat bervariasi ( d = 0,27–0,96, RMSE = 1–568 kg ha −1 ) ketika melihat rangkaian waktu pengukuran yang lengkap. Namun, itu lebih cocok dengan berat gabah akhir musim untuk sebagian besar eksperimen, meskipun eksperimen lain secara signifikan lebih tinggi atau lebih rendah dari prediksi. Waktu dan pengukuran berat gabah yang akurat, terutama di awal musim selama pembentukan gabah dan tahap awal pengisian gabah, sulit dilakukan selama eksperimen ini tetapi harus ditekankan untuk perbaikan model di masa mendatang. Secara umum, berat gabah yang disimulasikan dalam eksperimen dengan perlakuan pemupukan tinggi mendekati rata-rata nasional yang dilaporkan untuk semua kultivar benih sebesar 873 kg ha −1 (USDA NASS, 2024 ).
Biomassa akar selalu diprediksi berlebihan sebanyak tiga kali lipat dari nilai yang diamati di seluruh 13 percobaan dengan pengamatan. Model tersebut diharapkan untuk memprediksi berlebihan karena ekstraksi akar lengkap dalam percobaan lapangan hampir mustahil. Dari perspektif model, massa akar yang tinggi diperlukan untuk mempertahankan nutrisi tanaman yang cukup dan ketersediaan air dalam simulasi; oleh karena itu, prediksi berlebihan massa akar dianggap dapat diterima untuk versi model awal ini. Analisis yang lebih rinci dari simulasi individual disajikan dalam Bagian “ 3.2.2 . Pertumbuhan dan Perkembangan” untuk percobaan contoh untuk memamerkan simulasi pembagian biomassa, pembentukan benih, respons pupuk, dan kultivar yang berbeda. Semua pengamatan ditampilkan sebagai nilai rata-rata per tanggal dengan batang kesalahan satu standar deviasi untuk kejelasan visual, meskipun pengamatan individual dari tanggal yang sama sangat bervariasi karena variabilitas di lapangan yang besar. Keluaran grafis tambahan untuk semua simulasi individual, dengan titik data individual untuk pengukuran berulang, tersedia dalam Gambar S1-1 hingga S1-60 . Grafik-grafik ini juga menyertakan variabel keluaran yang tidak memiliki data teramati tetapi secara umum dipertimbangkan selama proses pengembangan model untuk memeriksa kewajarannya.
3.2.2 Pertumbuhan dan perkembangan
Untuk memverifikasi keakuratan model untuk akumulasi dan pembagian biomassa, efek N, dan perbedaan kultivar, pertumbuhan dan perkembangan simulasi contoh dianalisis lebih lanjut secara terperinci. Pembagian vegetatif (Gambar 4a ) umumnya diprediksi dengan baik kecuali untuk berat akar. Selama pembentukan tanaman, pertumbuhan biomassa terutama didorong oleh peningkatan berat daun. Dengan dimulainya pembungaan, pertumbuhan biomassa semakin bergeser ke arah berat batang dan biji. Peningkatan berat daun, dan kemudian juga berat batang, semakin mendatar karena asimilat bergeser ke arah pertumbuhan biji reproduktif dan tanaman mendekati kematangan fisiologis. Setelah kematangan fisiologis, penuaan daun meningkat dan pertumbuhan daun berakhir, yang mengakibatkan penurunan tajam pada berat daun hingga akhir simulasi. Perlambatan serupa dalam pertumbuhan, pendataran, dan sedikit penuaan akhir musim terjadi pada berat batang, meskipun berat batang secara keseluruhan lebih tinggi daripada berat daun. Penurunan berat akar yang terus-menerus disimulasikan selama paruh kedua musim tanam didorong oleh peningkatan mobilisasi N dan parameter penuaan untuk mewakili penjarangan sendiri dan penuaan tanaman jantan. Bahasa Indonesia: Dengan tidak adanya pengukuran berat akar yang akurat dari uji coba, dinamika berat akar dianggap dapat diterima untuk versi model awal tetapi harus dianalisis secara lebih rinci dalam uji coba mendatang. Jumlah simpul (Gambar 4b ) meningkat secara linear dari 0 hingga 20 dalam waktu sekitar 2,5 bulan setelah penanaman pada bulan Mei dan kemudian mencapai dataran tinggi pada akhir Juli hingga akhir simulasi. Ini cocok dengan pengamatan karena tanaman menghentikan pemanjangan dan penambahan simpul lebih lanjut dalam waktu 6 minggu setelah dimulainya pembungaan pada awal Juni. Pembentukan benih (Gambar 4c ) dimulai dengan dimulainya pembungaan, tetapi pengukuran pertama tidak dilakukan sampai akhir pembungaan pada awal Agustus. Benih mulai terisi dan terus bertambah berat dan dataran tinggi setelah sekitar 2 bulan pada akhir September. Simulasi ini cocok dengan berat benih akhir musim dengan baik, tetapi melebih-lebihkan perkembangan benih dalam musim. Ini dapat diterima dan lebih disukai dibandingkan dengan prediksi berat benih yang kurang. Pengukuran berat biji-bijian kemungkinan diremehkan karena biji-bijian hancur, dimakan burung, dan tantangan dalam memisahkan biji sepenuhnya dari batang dan daun untuk pengukuran, terutama untuk biji-bijian kecil selama pertumbuhan reproduksi awal. Perbandingan kultivar (Gambar 4d ) menunjukkan pola pertumbuhan biomassa yang berbeda dari kultivar IH Williams dan NWG 2730 dengan periode pertumbuhan yang lebih panjang secara keseluruhan dan biomassa puncak tiga kali lebih tinggi untuk IH Williams. Biomassa akhir musim untuk IH Williams sedikit lebih tinggi dari yang diperkirakan sejalan dengan perkiraan berat biji yang lebih tinggi, tetapi masih menangkap tren keseluruhan pengurangan biomassa karena penuaan daun dan tanaman jantan.

3.2.3 Respon Nitrogen
Total biomassa di atas permukaan tanah yang disimulasikan untuk IH Williams pada tahun 2022 adalah tiga hingga empat kali lebih tinggi untuk perlakuan 280 kg N ha −1 dibandingkan dengan perlakuan 0 kg N ha −1 (Gambar 5a ), yang sesuai dengan perbedaan yang diamati yang ditemukan dalam uji coba lapangan. Peningkatan stres N di bawah uji coba 0 kg N ha −1 juga mengurangi pemanjangan ruas dan tinggi tajuk keseluruhan (Gambar 5b ). Simulasi tersebut sangat cocok dengan perbedaan tinggi tajuk yang diamati kecuali pada bulan Juni, yang harus dianalisis lebih lanjut. Perlakuan N yang berbeda juga memengaruhi komposisi N jaringan yang disimulasikan seperti yang diharapkan (Gambar 5c,d ). Dimulai dengan konsentrasi N awal sebesar 5% di daun, konsentrasi tersebut tetap konstan atau sedikit meningkat pada perlakuan 280 kg N ha −1 hingga Juli, ketika permulaan pembentukan biji dan pengisian gabah serta penuaan daun di akhir musim menyebabkan mobilisasi N yang lebih tinggi dari jaringan daun dan selanjutnya pengurangan konsentrasi N daun menjadi 2,5% pada akhir musim. Untuk uji coba 0 kg N ha −1 , simulasi dimulai pada konsentrasi N daun yang sama sebesar 5%, tetapi dengan cepat berkurang menjadi 2,5%, diikuti oleh penurunan lebih lanjut menjadi 2% pada akhir musim karena stres N yang konsisten. Dinamika N jaringan batang mencerminkan pola jaringan daun tetapi pada tingkat yang lebih rendah secara keseluruhan karena komposisi protein yang lebih rendah untuk jaringan batang yang ditentukan untuk parameter model. Dinamika N jaringan yang disimulasikan secara keseluruhan dalam perlakuan tanpa N dan tinggi tampak wajar berdasarkan perbandingan dengan dinamika N jaringan pada tanaman lain dan juga sejalan dengan pengukuran N daun terpilih dari percobaan lapangan.
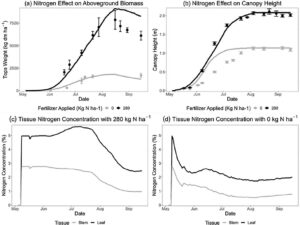
Proses pengembangan model iteratif mempertimbangkan data eksperimen dan wawasan dari literatur untuk mencapai simulasi pertumbuhan dan perkembangan rami berdasarkan karakteristik fisiologis rami dalam kerangka kerja CROPGRO umum. Hasil keseluruhan memuaskan untuk model awal ini untuk spesies baru. Perubahan dan perbaikan membuat model tersebut cocok untuk aplikasi lebih lanjut sambil menguraikan eksperimen mendatang, kebutuhan data, dan aspek kerangka kerja model yang dapat ditingkatkan.
3.3 Aplikasi model: Analisis tanggal tanam
Analisis musiman mengevaluasi variabilitas produksi rami di bawah tanggal penanaman variabel dari 1 Maret hingga 30 Juni dalam peningkatan 15 hari selama rentang waktu 21 tahun dari 2003 hingga 2023 di lokasi PSREU (Gambar 6 ) dan WFREC (Gambar 7 ). Tanpa mempertimbangkan efek tanggal penanaman, perbedaan simulasi umum antara kultivar IH Williams dan NWG 2730 dan dua lokasi tersebut sebanding dengan pengamatan dari percobaan lapangan. Produksi rami yang disimulasikan di lokasi WFREC mencapai biomassa yang lebih tinggi pada pembungaan dan kematangan gabah dibandingkan dengan rami yang disimulasikan di lokasi PSREU, sementara kultivar IH Williams mencapai biomassa yang lebih tinggi tetapi hasil gabah yang lebih rendah dibandingkan dengan kultivar NWG 2730. Durasi musim tanam berkisar antara 42 dan 58 hari dari penanaman hingga pembungaan dan 127 dan 144 hari dari penanaman hingga kematangan gabah untuk IH Williams. Untuk NWG 2730, musim tanam jauh lebih pendek dengan 34–52 hari dari penanaman hingga pembungaan dan 85–105 hari dari penanaman hingga kematangan biji-bijian. Untuk kultivar dan jenis panen, musim tanam umumnya lebih pendek untuk lokasi PSREU, mirip dengan yang diamati dalam percobaan lapangan. Untuk kultivar, lokasi, dan tanggal penanaman yang sama, variabilitas di antara 21 musim cukup sederhana karena pola cuaca yang umumnya stabil. Peristiwa cuaca abnormal seperti badai, kekeringan, hujan lebat, atau genangan air adalah efek abiotik yang terjadi selama uji coba eksperimental dan dapat mengurangi viabilitas tanaman secara signifikan dalam beberapa tahun. Namun, hal ini tidak dipertimbangkan dalam analisis ini atau dielakkan dalam simulasi dengan menginisialisasi tanah dengan kadar air awal yang sederhana dan irigasi atas selama pembentukan tanaman. Waktu panen dianalisis secara lebih rinci, pertama untuk tujuan serat di sekitar puncak pembungaan dan kedua untuk tujuan benih dan biomassa umum setelah pematangan biji-bijian. Tanggal penanaman memengaruhi periode pertumbuhan rata-rata untuk kedua jenis panen, dengan penanaman dari pertengahan Mei hingga pertengahan Juni memiliki periode pertumbuhan yang lebih panjang karena kondisi hari yang panjang setelah penanaman memperpanjang waktu hingga berbunga. Penanaman akhir Juni terjadi setelah hari dengan panjang hari terpanjang (titik balik matahari musim panas). Oleh karena itu, pertumbuhan terjadi pada panjang hari yang lebih pendek, sehingga menghasilkan waktu yang lebih pendek hingga berbunga. Efek umum dari tanggal penanaman pada waktu berbunga lebih kuat di lokasi WFREC yang lebih utara dan khususnya diekspresikan untuk kultivar IH Williams. Hal ini diharapkan karena efek fotoperiode yang lebih kuat (PPSEN) yang ditetapkan untuk kultivar ini. Untuk panen saat berbunga, hal ini menyebabkan perpanjangan periode pertumbuhan hingga 13 hari untuk IH Williams di lokasi WFREC saat membandingkan penanaman pada tanggal 1 Juni hingga 15 April (Gambar 7a)). Waktu pembungaan yang lebih lama juga disimulasikan untuk penanaman yang sangat awal di bulan Maret karena suhu yang lebih rendah memperlambat perkembangan sebagaimana didefinisikan dalam respons suhu kardinal CROPGRO-Hemp, bahkan dalam kondisi hari pendek. Beginilah cara kerja model dengan rangkaian parameter saat ini, tetapi harus dievaluasi lebih lanjut dalam uji coba khusus untuk menentukan fenologi dan waktu pembungaan dalam kondisi suhu rendah.
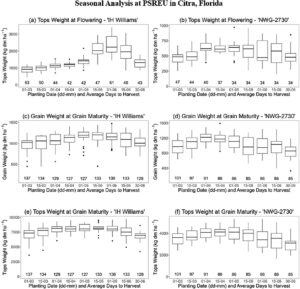
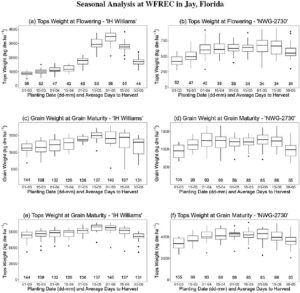
Sejalan dengan waktu yang diperpanjang untuk pembungaan untuk penanaman dari pertengahan Mei hingga pertengahan Juni, berat biomassa saat pembungaan secara signifikan lebih tinggi dibandingkan dengan penanaman lebih awal atau lebih lambat karena periode pertumbuhan vegetatif yang diperpanjang di bawah suhu dan radiasi tinggi. Mirip dengan efek tanggal penanaman pada waktu pembungaan, efek pada produktivitas biomassa lebih kuat di situs WFREC yang lebih utara dan khususnya diekspresikan untuk kultivar IH Williams. Misalnya, IH Williams di situs WFREC mencapai 3500 kg ha −1 biomassa rata-rata saat pembungaan untuk penanaman 1 Juni, dibandingkan dengan 1800 kg ha −1 untuk penanaman 1 Mei atau 30 Juni dan serendah 1000 kg ha −1 untuk penanaman pada bulan Maret. Sebaliknya, biomassa rata-rata untuk IH Williams saat pembungaan di situs PSREU secara signifikan lebih rendah untuk tanggal penanaman yang sama dan berkisar antara sekitar 1000 dan 2300 kg ha −1 . Penanaman awal pada bulan Maret tidak menghasilkan biomassa yang lebih besar meskipun periode pertumbuhan lebih lama hingga berbunga karena suhu dan radiasi yang lebih rendah, yang mengurangi fotosintesis dan mengasimilasi produksi. Untuk panen kedua, pada saat kematangan gabah untuk tujuan gabah atau biomassa keseluruhan, dampak tanggal penanaman kurang jelas. Untuk lokasi PSREU (Gambar 6c–f ), berat gabah rata-rata berkisar antara 1000 dan 1200 kg ha −1 untuk IH Williams dan 800 dan 1200 kg ha −1 untuk NWG 2730. Biomassa rata-rata berkisar antara 6000 dan 7500 kg ha −1 untuk IH Williams dan 3000 dan 4000 kg ha −1 untuk NWG 2730. Tidak ada pola yang jelas antara tanggal penanaman dan panen pada saat kematangan dapat disimpulkan dari grafik untuk lokasi ini. Untuk lokasi WFREC (Gambar 7c–f), hasilnya serupa tetapi dengan bobot gabah dan biomassa yang lebih tinggi secara keseluruhan sekitar 10%–30% dibandingkan dengan lokasi PSREU. Singkatnya, tanggal tanam tampaknya memiliki dampak yang kurang signifikan pada produksi gabah dan biomassa untuk panen pada saat gabah matang. Perbedaan dalam tingkat produksi yang disimulasikan tampaknya sebagian besar didorong oleh perbedaan umum dalam pola cuaca selama bagian awal atau akhir musim. Analisis lebih lanjut tentang mekanisme untuk simulasi pembentukan dan pematangan benih, seperti suhu kardinal yang ditetapkan sebelumnya, direkomendasikan untuk memverifikasi hasil ini. Oleh karena itu, sebagai hasil utama dari analisis musiman pertama ini, kami sarankan untuk berfokus pada dampak tanggal tanam pada fenologi dan produksi biomassa untuk panen saat berbunga untuk tujuan serat. Penanaman pada tanggal 1 Juni atau setidaknya antara tanggal 15 Mei dan 15 Juni optimal untuk biomassa tinggi saat berbunga. Ini mirip dengan kedelai di Florida utara, di mana tanggal tanam optimal adalah sekitar tanggal 1 Juni, dan penanaman lebih awal atau lebih lambat akan menghasilkan tanaman yang lebih pendek dengan biomassa dan hasil yang lebih sedikit. Relevansi hasil ini untuk produksi rami praktis harus didiskusikan lebih lanjut dengan para praktisi. Penanaman yang sangat awal dan terlambat dapat menimbulkan risiko pembekuan yang lebih tinggi pada awal atau akhir periode pertumbuhan, khususnya di lokasi WFREC yang lebih utara, atau dapat berbenturan dengan rotasi tanaman yang ada. Memindahkan tanggal penanaman ke akhir Mei atau awal Juni juga meningkatkan risiko hujan yang lebih lebat dan lebih sering, yang merupakan ciri khas musim panas di Florida, yang dapat membuat ladang tidak cocok untuk ditanami.
Analisis musiman sekunder difokuskan pada produksi kultivar IH Williams di lokasi PSREU dan WFREC dan panen pada saat berbunga dengan waktu yang lebih lama hingga berbunga karena hal ini diidentifikasi sebagai masalah manajemen kritis untuk produksi rami (Gambar 8 ). Perlakuan simulasi dengan CSDL yang dikurangi (genetik) atau panjang hari yang diperpanjang (pencahayaan) dimaksudkan untuk mewakili kemajuan pemuliaan hipotetis atau skenario manajemen dengan pencahayaan intensitas rendah tambahan untuk mengatasi kondisi hari pendek atau produksi di lintang yang lebih utara dengan iklim yang identik. Dibandingkan dengan perlakuan “dasar” dengan panjang hari yang diamati dan CSDL genetik 13,45 jam, semua perlakuan “genetik” memperpanjang periode pertumbuhan vegetatif dan dengan demikian meningkatkan biomassa simulasi pada titik pembungaan. Hal yang sama terjadi dengan perpanjangan panjang hari buatan. Peningkatan biomassa simulasi pada saat pembungaan berkorelasi dengan kekuatan perlakuan. Peningkatan panjang hari sebesar 2,0 jam menyebabkan peningkatan biomassa yang lebih kuat pada saat pembungaan dibandingkan dengan peningkatan panjang hari sebesar 1,0 atau 0,5 jam. Pengurangan panjang hari pendek kritis (CSDL) dari kultivar sebanyak 1,0 jam menyebabkan peningkatan biomassa yang lebih kuat saat berbunga dibandingkan dengan pengurangan CSDL sebanyak 0,5 atau 0,25 jam. Untuk penanaman dari Mei hingga Juni, yang diidentifikasi sebagai yang paling produktif dalam analisis sebelumnya, perlakuan yang sudah sederhana seperti peningkatan panjang hari 0,5 jam atau pengurangan CSDL 0,25 jam menunjukkan peningkatan yang signifikan dalam berat biomassa saat berbunga dengan faktor sekitar 2–4. Hal ini menghasilkan total biomassa saat berbunga dalam kisaran 8000–12.000 kg ha −1 , sebanding dengan uji coba penelitian hasil tinggi dari kultivar yang sama di garis lintang utara di Amerika Serikat (University of Illinois, 2024 ). Penanaman lebih awal pada bulan April dan Maret akan memerlukan perpanjangan panjang hari yang lebih besar untuk mencapai produktivitas yang sama saat berbunga, dengan penanaman pada tanggal 1 Maret hanya mencapai peningkatan yang berarti dalam produktivitas biomassa di bawah perpanjangan siang hari maksimum 2,0 jam. Kombinasi perlakuan dan tanggal tanam individual, seperti panjang hari +1 jam dan CSDL -1,0 untuk penanaman tanggal 15 Maret di lokasi WFREC, menunjukkan variabilitas tinggi di antara 21 musim. Ini menunjukkan bahwa efek suhu rendah pada fenologi berkontribusi pada perpanjangan waktu pembungaan dan peningkatan pertumbuhan vegetatif, tetapi pada tingkat yang bervariasi dari musim ke musim. Sebagai kesimpulan, hasil simulasi mengungkapkan potensi yang signifikan untuk meningkatkan produksi biomassa saat pembungaan untuk kultivar IH Williams dengan kultivar hipotetis dan tanggal tanam yang lebih baik. Peningkatan tersebut mungkin terjadi untuk kedua lokasi, PSREU di Citra, FL, dan WFREC di Jay, FL, tetapi tergantung pada tanggal dan lokasi penanaman, jenis dan besarnya perlakuan yang diperlukan untuk mencapai peningkatan produktivitas tertentu bervariasi.
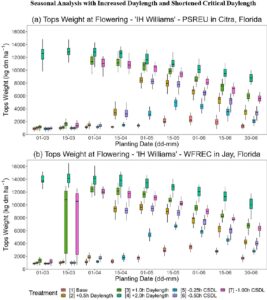
3.4 Keterbatasan
Kekuatan analitis dari studi ini dibatasi oleh pendekatan metodologisnya, fokus geografis pada Florida, serta kualitas dan keragaman data eksperimen yang digunakan. Memilih CROPGRO sebagai templat pemodelan, dan pemodelan berbasis proses sebagai metode secara umum, memiliki banyak keuntungan tetapi juga beberapa keterbatasan intrinsik (KJ Boote et al., 1996 ; Jones et al., 2017 ; Spitters, 1990 ). Model berbasis proses memperkirakan sistem pertanian yang kompleks karena keterbatasan komputasi atau berbagai tingkat pemahaman tentang komponen sistem individual. Model-model ini juga bergantung pada data yang luas dan berkualitas tinggi, tetapi ruang lingkup dan intensitas pengumpulan data dibatasi oleh sumber daya keuangan dan manusia yang tersedia. Lebih jauh lagi, tantangan budidaya dan pengelolaan umum adalah hal yang umum ketika melakukan uji coba lapangan untuk tanaman baru dengan riwayat budidaya lokal yang terbatas atau tidak ada, yang mungkin memiliki bias pengumpulan data. Misalnya, heterogenitas kinerja tanaman di berbagai area lahan karena variasi tanah lokal yang mungkin tidak diketahui, kepadatan penanaman yang berbeda, atau variabilitas benih internal dapat memengaruhi pengukuran tetapi tidak termasuk dalam kerangka CROPGRO, yang mengasumsikan kondisi pertumbuhan dan kinerja tanaman yang homogen di seluruh area simulasi. Terakhir, hasil penerapan model didasarkan pada simulasi dan perlu evaluasi lebih lanjut dengan data eksperimen yang diamati. Beberapa kemungkinan untuk perbaikan model dan penelitian di masa mendatang diidentifikasi selama proses pengembangan model.
3.5 Penelitian masa depan
Penelitian di masa depan harus mencakup eksperimen tambahan dan evaluasi model untuk lokasi, lingkungan, dan kultivar lain, termasuk kultivar khusus seperti rami berbatang kuning dengan sifat fisiologis yang unik (Blandinières et al., 2023 ), untuk membuat model lebih kuat. Secara khusus, pengukuran yang lebih tepat dan luas pada fenologi di berbagai garis lintang dan tanggal penanaman, serta pengamatan terperinci tentang pembentukan dan pematangan benih, akan berharga untuk peningkatan model lebih lanjut. Di luar prediksi biomassa saat ini, simulasi karakteristik kualitas akan menjadi tambahan penting bagi model untuk studi masa depan. Parameter kualitas serat sebagian ditentukan sebelumnya melalui genetika, seperti partisi menjadi serat kulit pohon dan hurd pada batang rami, tetapi penelitian lebih lanjut diperlukan pada dampak lingkungan yang potensial (Salentijn et al., 2015 ). Ini juga meluas ke pemodelan pasca panen, seperti Liu et al. ( 2015 ) menemukan bahwa pengaturan waktu panen dan perendaman lapangan, proses pasca panen terpisah yang bergantung pada cuaca, memiliki dampak signifikan pada parameter kualitas serat dan perlu dikelola secara sistematis. Untuk benih rami, konsentrasi minyak dan protein merupakan kriteria penting yang dipengaruhi oleh interaksi genotipe × lingkungan (Irakli et al., 2019 ; Lan et al., 2019 ). Banyak model gandum ( Triticum aestivum ) (Asseng et al., 2018 ) dan CROPGRO menyediakan subrutin untuk simulasi protein dan minyak benih. Produksi bunga atau trikoma kelenjar resin dan kanabinoid tidak tertangkap dalam model awal ini yang diadaptasi untuk aplikasi serat dan benih, di mana representasi pembungaan hanya disertakan sebagai peristiwa fenologis. Konsentrasi kanabinoid di sebagian besar kultivar serat dan benih sangat rendah dan tidak dimaksudkan sebagai produk panen. Namun, hal ini dapat relevan untuk kepatuhan hukum, potensi penggunaan produk sampingan, atau sistem penanaman penggunaan ganda, dan simulasi konsentrasi kanabinoid harus dipertimbangkan untuk pengembangan model di masa mendatang. Hal ini dapat mengarah pada adaptasi lebih lanjut dari model untuk simulasi bunga rami yang kaya kanabinoid, yang akan memerlukan representasi bunga dan trikoma sebagai organ tanaman. Hal ini sebanding dengan adaptasi CROPGRO baru-baru ini terhadap bunga kesumba ( Carthamus tinctorius L.) oleh Singh et al. ( 2016 ) dan dapat dikombinasikan dengan pendekatan pemodelan untuk pengembangan kanabinoid selama pertumbuhan dan perkembangan, seperti yang dianalisis oleh R. Yang et al. ( 2020)). Selain itu, kerja partisipatif di masa mendatang dengan para pemangku kepentingan dapat mengembangkan model tanaman ini menjadi alat pendukung keputusan yang lebih dapat ditindaklanjuti atau program edukasi yang dapat digunakan oleh petani dan praktisi untuk mengeksplorasi interaksi genotipe × lingkungan × pengelolaan dan mengembangkan pendekatan untuk budidaya rami yang berkelanjutan. Ini akan menjadi langkah penting untuk membuat model tanaman lebih relevan bagi pengguna dan meningkatkan adopsi model dengan mempertimbangkan potensi penggunaan akhir (KJ Boote et al., 1996 ; Cerf et al., 2012 ; Prost et al., 2012 ). Terakhir, akan bermanfaat untuk melakukan perbandingan model sistematis dari model rami yang awalnya ditinjau menggunakan kumpulan data umum mengikuti contoh inisiatif Agricultural Model Intercomparison and Improvement Project (AgMIP) (Rosenzweig et al., 2013 ).
4 KESIMPULAN
Studi ini mengadaptasi dan mengkalibrasi model tanaman CROPGRO-Hemp baru berdasarkan informasi literatur dan pengumpulan serangkaian data eksperimen awal dan menghasilkan simulasi akurat pertumbuhan dan perkembangan produksi serat dan biji rami di Florida. Analisis tanggal tanam musiman menunjukkan penerapan model untuk menunjukkan variabilitas biomassa musiman yang disimulasikan dan produktivitas biji-bijian karena panjang hari dan efek suhu selama 21 musim tanam di dua lokasi. Lebih jauh, analisis ini mengungkapkan dampak signifikan dari pilihan tanggal tanam pada produktivitas biomassa karena dampak pada panjang hari dan fenologi. Selain itu, analisis ini mengevaluasi potensi produksi dengan perpanjangan panjang hari hipotetis atau kultivar baru yang memiliki panjang hari pendek kritis yang lebih pendek. Studi lebih lanjut dan penyesuaian parameter model akan berharga untuk meningkatkan simulasi pembagian vegetatif, pembungaan yang tepat, dan fenologi pembentukan biji dan untuk memverifikasi model untuk varietas lain dan lokasi tumbuh, khususnya untuk bunga rami. Rilis model berbasis proses yang tersedia untuk umum dan mudah diakses ini merupakan tambahan yang berharga bagi komunitas rami industri dan bidang pemodelan tanaman secara umum.



Tinggalkan Balasan