Keranjang Anda kosong!
Analisis perbandingan metabolit primer dan sekunder pada bibit gandum ( Triticum aestivum L.) yang dibudidayakan dengan kepadatan fluks foton fotosintesis dan periode pertumbuhan yang berbeda

Abstrak
LATAR BELAKANG
Untuk menjelaskan dampak faktor lingkungan terhadap kualitas nutrisi metabolit pada bibit gandum, baik metabolit primer maupun sekunder harus diperiksa secara komprehensif. Oleh karena itu, kami melakukan analisis komparatif metabolit ini pada bibit gandum yang tumbuh dalam berbagai kerapatan fluks foton fotosintesis (PPFD) (200, 400, dan 800 μmol m −2 s −1 ) dan periode pertumbuhan (5, 7, 9, 11, 13, dan 15 hari) untuk membantu mengoptimalkan kualitas nutrisi bibit.
HASIL
Secara total, 74 metabolit dikarakterisasi, termasuk 51 metabolit primer dan 23 metabolit sekunder. Analisis multivariat mengonfirmasi perubahan flavonoid, yang merupakan antioksidan yang diketahui, dalam berbagai kondisi PPFD. Bibit gandum yang tumbuh pada PPFD 400 μmol m −2 s −1 selama lebih dari 9 hari menunjukkan kandungan flavonoid yang lebih tinggi daripada yang tumbuh dalam kondisi lain. Lebih jauh, peningkatan metabolisme sistein dan metionina, metabolisme gula, dan biosintesis karotenoid dan fenilpropanoid diamati.
KESIMPULAN
PPFD sebesar 400 μmol m −2 s −1 meningkatkan metabolisme gula pada bibit gandum dengan mendorong anabolisme melalui fotosintesis. Lebih jauh lagi, akumulasi gula, bahan baku utama untuk metabolit sekunder, memengaruhi biosintesis flavonoid. Hasil-hasil ini menunjukkan bahwa bibit gandum menjalani pemrograman ulang metabolik sebagai respons terhadap intensitas cahaya, yang memungkinkan respons adaptif. Dengan demikian, penelitian kami mengonfirmasi hubungan antara metabolit primer dan sekunder pada bibit gandum dalam berbagai kondisi PPFD. Lebih jauh lagi, wawasan yang diperoleh meningkatkan pemahaman kita tentang pengaruh intensitas dan durasi cahaya pada akumulasi metabolit, yang sangat penting untuk mengoptimalkan praktik pertanian. © 2025 Penulis. Jurnal Ilmu Pangan dan Pertanian diterbitkan oleh John Wiley & Sons Ltd atas nama Society of Chemical Industry.
PERKENALAN
Bibit gandum telah menarik perhatian sebagai makanan kesehatan karena tingginya kadar vitamin, makronutrien, enzim, dan asam amino. 1 , 2 Budidaya bibit gandum memungkinkan kontrol yang tepat dari berbagai parameter pertumbuhan seperti saturasi air, suhu, cahaya, kelembaban dan periode pertumbuhan. 3 Intensitas dan kualitas cahaya secara nyata mempengaruhi metabolisme tanaman, termasuk fotosintesis, produksi gula, dan biosintesis dan akumulasi metabolit sekunder. 4 , 5 Oleh karena itu, memahami efek kerapatan fluks foton fotosintesis (PPFD) sangat penting untuk mengoptimalkan kondisi pertumbuhan tanaman. PPFD mengacu pada jumlah foton yang mencapai area 1 m 2 per detik, dan pengukuran parameter ini memungkinkan kontrol intensitas cahaya yang tepat. 4 – 6 Bibit gandum memerlukan tingkat intensitas cahaya tertentu pada tahap pertumbuhan yang berbeda untuk memaksimalkan fotosintesis dan perkembangannya. 7 Pada gandum, intensitas cahaya secara selektif memengaruhi distribusi dan redistribusi zat gizi makro dan mikro 8 dan, pada kecambah dan mikrohijauan, akumulasi metabolit sekunder yang meningkatkan kesehatan berbeda-beda di antara kondisi PPFD. 9 Bibit gandum menunjukkan sifat antioksidan, yang meningkat secara substansial selama perkecambahan, terutama disebabkan oleh pembentukan flavonoid, sejenis metabolit sekunder. 10 Dengan demikian, perubahan metabolik pada bibit gandum sebagai respons terhadap PPFD perlu dijelaskan untuk membantu mengoptimalkan produksi bahan-bahan yang meningkatkan kesehatan; namun, aspek ini sebagian besar masih belum dieksplorasi.
Metabolomik berfungsi sebagai alat yang berguna untuk mendeteksi respons tanaman terhadap perubahan lingkungan dan menjelaskan mekanisme biologis yang mendasarinya. 11 – 13 Lebih jauh lagi, pemahaman yang komprehensif tentang jalur biosintesis yang menghubungkan metabolit primer dan sekunder diperlukan, mengingat bahwa metabolit primer adalah prekursor metabolit sekunder. 14 Dalam Arabidopsis, intensitas cahaya yang berbeda mengakibatkan perbedaan gula, perantara siklus asam trikarboksilat (TCA), dan kandungan asam lemak 15 ; gula lebih melimpah di Arabidopsis yang tumbuh dalam kondisi cahaya tinggi (500–600 μmol m −2 s −1 ), sedangkan alanin, glutamin, asam glutamat, dan asam aspartat lebih melimpah pada tanaman yang tumbuh dalam kondisi cahaya pertumbuhan normal (130 μmol m −2 s −1 ). 15 Intensitas cahaya yang tinggi umumnya mendorong produksi metabolit sekunder, sedangkan pada beberapa tanaman seperti skiofit, termasuk ginseng, produksi metabolit sekunder didorong pada intensitas cahaya rendah. Dengan demikian, kondisi cahaya yang optimal untuk pertumbuhan dan perkembangan tanaman berbeda di antara taksa. 16
Studi kami sebelumnya mengonfirmasi bahwa perbedaan senyawa fenolik pada bibit gandum lebih menonjol antar periode pertumbuhan daripada antar kultivar. 17 Hal ini menunjukkan bahwa faktor lingkungan memainkan peran penting dalam menentukan komposisi nutrisi bibit gandum. Namun, hanya sedikit studi yang mengeksplorasi jaringan metabolisme kompleks melalui profil metabolit primer dan sekunder pada bibit gandum. Studi saat ini dilakukan untuk menyelidiki kualitas nutrisi bibit gandum melalui analisis komprehensif terhadap perubahan metabolit primer dan sekunder sebagai respons terhadap berbagai tingkat PPFD dan periode pertumbuhan. Untuk tujuan ini, kami membudidayakan bibit gandum menggunakan berbagai kondisi PPFD (200, 400, dan 800 μmol m −2 s −1 ) dan periode pertumbuhan (5, 7, 9, 11, 13, dan 15 hari). Selanjutnya, profil metabolit primer dan sekunder berdasarkan kromatografi gas (GC) dan kromatografi cair-spektrometri massa (MS) dilakukan. Perubahan metabolit bibit gandum menurut faktor lingkungan diselidiki menggunakan analisis multivariat. Hasil penelitian ini diharapkan dapat meningkatkan pemahaman umum tentang parameter budidaya tanaman secara signifikan untuk mengoptimalkan produksi.
BAHAN DAN METODE
Bahan kimia dan reagen
Sigma-Aldrich (St Louis, MO, AS) memasok ribitol, metoksiamina klorida, asam askorbat, 6-metoksiflavon, piridina, campuran metil ester asam lemak (FAME) (C8–C24), 5α-kolestana, asam pentadekanoat, dan N -metil- N- trimetilsilil trifluoroasetamida (MSTFA). Metanol dan etanol diperoleh dari Daejung (Kyungki-do, Republik Korea), sedangkan kloroform bersumber dari Burdick dan Jackson (Muskegon, MI, AS). Thermo Fisher Scientific (Waltham, MA, AS) memasok asam klorida, toluena, dan heksana, dan kalium hidroksida (KOH) diperoleh dari Wako (Wako Pure Chemical Corporation, Osaka, Jepang). Thermo Fisher Scientific juga menyediakan metoksiamina hidroklorida (MOX), dan trans – β- apo-8-karotenal diperoleh dari CaroteNature (Münsingen, Swiss). Kecuali dinyatakan lain, semua senyawa yang digunakan dalam penelitian ini bermutu HPLC.
Persiapan sampel
Kultivar gandum Korea Saekeumkang digunakan dalam penelitian ini. Ditanam pada tahun 2023 dengan tanah buatan dalam kondisi ruang pertumbuhan yang terkendali. Kondisi pertumbuhan untuk semua tanaman dipertahankan pada kelembaban relatif 60–70%, suhu 18–22 °C, dan siklus foto hari pendek 9 jam terang dan 15 jam gelap. Tanaman mengalami kondisi PPFD yang berbeda, meliputi 200, 400, dan 800 μmol m −2 s −1 , berdasarkan kisaran intensitas cahaya khas yang diamati dalam kondisi pengolahan tanah (PPFD 150–900 μmol m −2 s −1 ). 7 , 8 Kondisi ini dipilih untuk mewakili intensitas cahaya rendah hingga tinggi. Bibit gandum dipanen pada enam tahap pertumbuhan yang berbeda (5, 7, 9, 11, 13, dan 15 hari pasca perkecambahan) dan awalnya dikeringkan dengan udara di bawah cahaya alami dan suhu selama 3 hari pada suhu 25 °C. Setelah itu, sampel dikeringkan dengan beku pada suhu -78 °C dan disimpan pada suhu -80 °C hingga dilakukan analisis.
Ekstraksi dan analisis metabolit primer
Analisis zat hidrofilik (asam amino bebas, gula, alkohol gula, dan asam organik) dilakukan seperti yang dijelaskan sebelumnya. 18 Secara singkat, 10 mg sampel bibit gandum beku-kering yang digiling dicampur dengan 1 mL larutan metanol:air:kloroform (2,5:1:1, v:v:v) dalam tabung 2 mL. Sebagai standar internal (IS), 0,06 mL ribitol (0,2 mg mL −1 dalam metanol) ditambahkan. Setelah pengeringan, sampel pekat diderivatisasi dengan 0,08 mL MOX (20 mg mL −1 dalam piridina) dan kemudian diolah dengan 0,08 mL MSTFA. Metabolit hidrofilik diperiksa menggunakan GC-time-of-flight MS Benchtop (LECO, St Joseph, MI, AS) dan model 7890B GC (Agilent, Santa Clara, CA, AS) yang dipasangi kolom CP-SIL 8 CB (CP5860; Agilent); 1 μL sampel derivatisasi disuntikkan, dan mode split digunakan pada rasio 1:25. Suhu port injeksi, jalur transfer, dan sumber ion masing-masing ditetapkan pada 230, 250, dan 250 °C. Suhu oven dipertahankan pada 80 °C selama 2 menit, kemudian ditingkatkan pada laju 15 °C min −1 hingga 320 °C dan dipertahankan selama 10 menit. Data spektral dipindai pada rentang m / z 85–600. Luas puncak relatif dihitung sebagai rasio luas puncak relatif dibagi dengan luas puncak IS. 18
Ekstraksi asam lemak dari bibit gandum dilakukan dengan menggunakan metode yang telah dijelaskan sebelumnya. 19 Secara singkat, sampel bibit gandum yang dikeringkan beku (10 mg) ditempatkan dalam tabung 15-mL yang berisi 100 μL asam pentadekanoat (1 mg mL −1 dalam kloroform; digunakan sebagai IS) dan 2,5 mL larutan kloroform:metanol (2:1, v/v). Untuk metilasi, sampel pekat dicampur dengan 0,3 mL boron trifluorida 0,9 m dalam metanol. Asam lemak dianalisis menggunakan detektor ionisasi nyala model 7890B GC (Agilent) dengan kolom DB-WAX (122–7032, panjang 30 m, diameter 0,25 mm, ketebalan film 0,25 μm; Agilent). Nitrogen digunakan sebagai gas pembawa pada laju alir 1,00 mL min −1 . Suhu saluran masuk depan dan detektor adalah 250 °C. Suhu oven dipertahankan pada 130 °C selama 3 menit dan kemudian ditingkatkan pada laju 20 °C min -1 hingga 230 °C. Suhu akhir ditingkatkan pada laju 3 °C min -1 hingga 250 °C dan dipertahankan selama 5 menit. 19 Analisis kualitatif dan kuantitatif dilakukan dengan membandingkan rasio luas puncak sampel dengan rasio campuran FAME sebagai standar. 19
Ekstraksi klorofil dilakukan seperti yang dilaporkan sebelumnya 20 ; singkatnya, sampel bibit gandum beku-kering (10 mg) dicampur dengan 1 mL metanol. Campuran tersebut diaduk selama 30 detik dan kemudian disonikasi selama 30 menit pada suhu 70 °C menggunakan thermomixer (model 5355; Eppendorf AG, Hamburg, Jerman). Sampel yang disonikasi disentrifugasi pada 800 × g dan 4 °C selama 10 menit menggunakan konsentrator sentrifugal (CC-105; TOMY, Tokyo, Jepang). Spektrofotometer (Optizen POP; Mecasys Co., Daejeon, Republik Korea) digunakan untuk mengukur absorbansi supernatan pada 666 dan 653 nm. Total kandungan klorofil dihitung menggunakan rumus Wellburn. 20
Ekstraksi dan analisis metabolit sekunder
Zat lipofilik sekunder, seperti polikosanol, tokoferol, dan sterol, diekstraksi seperti yang dijelaskan sebelumnya. 21 Secara singkat, 10 mg bibit gandum beku-kering diekstraksi dengan 3 mL etanol asam askorbat 0,55 m dan 50 μL 5α-cholestane (10 μg mL −1 ) sebagai IS. Kemudian, 30 μL MSTFA dan 30 μL piridina ditambahkan untuk derivatisasi, diikuti dengan inkubasi pada suhu 60 °C selama 30 menit dalam thermomixer (model 5355; Eppendorf AG). Untuk pemisahan, alikuot 1-μL disuntikkan pada rasio split 1:10 ke dalam sistem GC-MS-QP2010 Ultra (Shimadzu, Kyoto, Jepang) yang dilengkapi dengan kolom Rtx-5MS (30 m × 0,25 mm, diameter dalam 0,25 μm; Restek). Gas helium dikirimkan pada laju tetap 1 mL min −1 . Suhu saluran masuk adalah 290 °C. Suhu oven awalnya 150 °C selama 2 menit, kemudian meningkat menjadi 320 °C pada laju 15 °C min −1 dan dipertahankan selama 10 menit. Suhu sumber ion dan kontak masing-masing adalah 230 dan 280 °C. Spektrum diperoleh pada rentang m / z 85–600 dan ion diidentifikasi dalam mode pemantauan ion terpilih untuk analisis puncak. Data kromatografi dianalisis menggunakan perangkat lunak larutan GCMS LabSolutions, versi 4.11 (Shimadzu). Kurva kalibrasi yang akurat untuk kuantifikasi absolut dibuat untuk setiap standar lipofilik pada beban mulai dari 0,025 μg hingga 5,00 μg, dengan bobot IS tetap 0,50 μg. 21
Prosedur ekstraksi dan penentuan karotenoid sedikit dimodifikasi dari metode yang dipublikasikan sebelumnya. 22 Secara singkat, sampel bibit gandum beku-kering (10 mg) dicampur dengan 3 mL asam askorbat 0,55 m dalam etanol dalam tabung 15 mL dan diaduk. Air (1,5 mL), heksana (0,75 mL), toluena (0,75 mL) dan 100 μL trans- β -apo-8-karotenal 25 ppm (sebagai IS) ditambahkan. Ekstrak pekat dilarutkan dalam 250 μL larutan metanol:diklorometana (1:1, v/v) dan disaring menggunakan filter spuit 0,5 μm. Karotenoid dipisahkan menggunakan sistem HPLC model 1100 (Agilent) yang dilengkapi dengan detektor dioda-array yang diatur pada 450 nm, pada kolom HPLC karotenoid YMC (250 × 4,6 mm, diameter dalam 3 μm) (YMC, Kyoto, Jepang). Kurva kalibrasi dibuat dengan membagi luas puncak IS dengan luas puncak senyawa standar untuk kuantifikasi. 22
Proses ekstraksi dan estimasi senyawa flavonoid sedikit dimodifikasi dari metode yang dijelaskan sebelumnya. 23 Kami menempatkan 50 mg bibit gandum dan 2 mL galangin (IS) 2,5 ppm dalam metanol dalam tabung 5 mL. Sampel yang difilter (10 μL) dianalisis menggunakan MS tandem dan detektor susunan fotodioda yang terhubung ke sistem kromatografi cair kinerja ultra (UPLC) (ACQUITY UPLC I-Class PLUS’ Waters, Milford, MA, AS). UPLC dipasangi kolom ACQUITY UPLC CSH C18 1,7 μm (150 × 2,1 mm, ketebalan kolom 1,7 μm; Waters). Kuantifikasi dilakukan menggunakan kurva kalibrasi yang dihasilkan dengan membagi luas puncak IS dengan luas puncak senyawa standar.
Analisis statistik
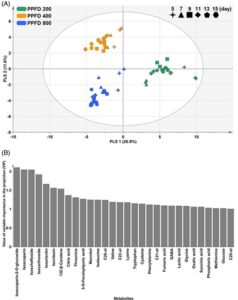
Setiap kelompok terdiri dari tiga replikasi biologis bibit gandum. Sebelum analisis multivariat, data disesuaikan menggunakan penskalaan varians unit. Analisis komponen utama (PCA), analisis diskriminan kuadrat terkecil parsial (PLS-DA) dan proyeksi ke struktur laten (PLS) dilakukan menggunakan perangkat lunak SIMCA, versi 14.1; Umetrics, Umeå, Swedia). Analisis pengayaan jalur berdasarkan basis data jalur Kyoto Encyclopedia of Genes and Genomes (KEGG) ( https://www.genome.jp/kegg ) dilakukan menggunakan MetaboAnalyst, versi 6.0 ( https://www.metaboanalyst.ca ; diakses pada 4 Maret 2024). Selain itu, analisis varians dilakukan menggunakan MetaboAnalyst, versi 6.0 (diakses 4 Maret 2024) untuk menentukan perbedaan signifikan [tingkat penemuan palsu (FDR) < 0,05] di antara bibit gandum yang ditanam dalam kondisi PPFD dan periode pertumbuhan yang berbeda. HASIL DAN PEMBAHASAN Profil metabolik bibit gandum Bibit gandum kaya akan metabolit sekunder yang terkait dengan sifat antioksidan, 23 dan, selama perkecambahan, faktor lingkungan memengaruhi konsentrasi metabolit ini. 2 Metabolit primer, yang memiliki sifat nutrisi yang berharga, terlibat langsung dalam pertumbuhan, perkembangan, dan reproduksi, berfungsi sebagai komponen metabolit sekunder. 19 Oleh karena itu, untuk memahami dampak faktor lingkungan terhadap kualitas nutrisi metabolit bibit gandum, analisis komprehensif terhadap metabolit primer dan sekunder sangat penting. Kami melakukan profil metabolik primer dan sekunder dari bibit gandum yang dibudidayakan dalam berbagai kondisi PPFD (200, 400 dan 800 μmol m −2 s −1 ) dan periode pertumbuhan (5, 7, 9, 11, 13 dan 15 hari) dan mengkarakterisasi total 74 metabolit, yang terdiri dari 51 metabolit primer, termasuk asam amino, asam lemak dan asam organik, dan 23 metabolit sekunder, seperti polikosanol, sterol, flavonoid dan karotenoid (lihat Informasi pendukung, Tabel S1–S18 ). Analisis multivariat bibit gandum yang dibudidayakan dengan kepadatan fluks foton fotosintesis dan periode pertumbuhan yang berbeda Analisis multivariat digunakan untuk menetapkan pentingnya faktor-faktor kunci di berbagai variabel dan mengungkap proses biokimia yang kompleks. 24 Untuk menilai dampak intensitas cahaya terkontrol dan periode pertumbuhan pada metabolisme bibit gandum, kami melakukan analisis multivariat, termasuk PCA, PLS-DA, pemodelan PLS dan analisis pengayaan jalur. Pertama, PCA dilakukan untuk menilai korelasi keseluruhan di antara sampel bibit gandum yang tumbuh dalam berbagai kondisi PPFD dan periode pertumbuhan (lihat Informasi pendukung, Gambar S1 ). Teknik statistik ini mengurangi sampel dari ruang berdimensi tinggi ke ruang berdimensi rendah sambil mempertahankan korelasi yang bermakna. 25 Dalam penelitian kami, model PCA mengungkapkan bahwa jumlah dari dua komponen utama (PC) teratas menyumbang 44,8% dari total varians (29,7% untuk PC1 dan 15,1% untuk PC2). PC1 dengan jelas membedakan bibit gandum yang dibudidayakan di bawah PPFD 200 μmol m −2 s −1 (PPFD 200) dari yang ditanam di bawah PPFD 400 μmol m −2 s −1 (PPFD 400) dan 800 μmol m −2 s −1 (PPFD 800). Mengenai periode pertumbuhan, bibit gandum yang ditanam selama 5 hari cenderung berbeda dari yang ditanam untuk periode pertumbuhan lainnya, sedangkan bibit gandum yang ditanam selama lebih dari 7 hari tidak menunjukkan pemisahan yang jelas. Hasil ini menunjukkan bahwa efek intensitas cahaya pada metabolisme bibit gandum lebih kuat daripada periode pertumbuhan. Pengaruh kerapatan fluks foton fotosintesis terhadap konsentrasi metabolit pada bibit gandum Berdasarkan hasil sebelumnya, PLS-DA dilakukan untuk memeriksa lebih lanjut perubahan metabolik pada bibit gandum menurut kondisi PPFD (Gbr. 1A ). PLS-DA adalah pendekatan statistik yang memutar PCA untuk mencapai pemisahan maksimum antara kelompok dalam kondisi tertentu. 26 Kami menggunakan kondisi PPFD sebagai variabel klasifikasi untuk membedakan model untuk PPFD 200, PPFD 400 dan PPFD 800. Kecocokan dan kemampuan prediktif model yang dihasilkan dievaluasi dari nilai R 2 dan Q 2 masing-masing. Model PLS-DA yang dihasilkan secara efektif mewakili perbedaan dalam sampel berdasarkan kondisi cahaya, dengan nilai R 2 dan Q 2 yang tinggi (> 0,5). Untuk mengidentifikasi metabolit utama yang berkontribusi pada pemisahan ini, variabel penting dalam plot proyeksi (VIP) dari masing-masing model PLS-DA diproduksi (Gbr. 1B ). Dalam plot VIP, metabolit dengan VIP > 1 dianggap berkontribusi secara substansial pada pemisahan sampel. 26 Dalam penelitian ini, flavonoid seperti isoscoparine-2- O -glucoside, isoscoparine, isochaftoside, isocarinoside, isoorientin dan isovitexin menunjukkan nilai VIP tertinggi. Penelitian sebelumnya mengkarakterisasi profil metabolit fenolik dan kapasitas antioksidan dari kultivar bibit gandum dan periode pertumbuhan. 23 Bibit gandum merupakan sumber flavonoid yang kaya dan bermanfaat bagi kesehatan manusia. Sejauh pengetahuan kami, sejauh ini belum ada penelitian yang melaporkan perubahan metabolik pada bibit gandum yang dipengaruhi oleh kondisi PPFD. Oleh karena itu, hasil penelitian ini mengonfirmasi, untuk pertama kalinya, bahwa perubahan PPFD berdampak signifikan pada metabolisme flavonoid pada bibit gandum.
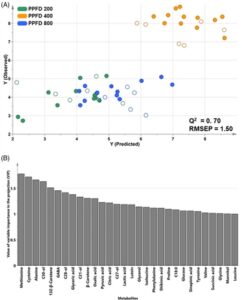
Metabolit sekunder tanaman berasal dari metabolit primer, sehingga pemahaman yang komprehensif tentang jalur biosintesis metabolit primer diperlukan. 19 Namun, interpretasi jaringan metabolisme kompleks pada bibit gandum melalui profil metabolit primer dan sekunder masih kurang. Oleh karena itu, model PLS prediktif berdasarkan hubungan antara metabolit primer dan sekunder pada bibit gandum telah disesuaikan (Gbr. 2 ). PLS melibatkan ekstraksi variabel baru yang memaksimalkan kovariansi antara kombinasi linear variabel independen ( X ) dan variabel dependen ( Y ). 21 Untuk pengembangan model, sampel dibagi menjadi dua kelompok: set pelatihan, yang digunakan untuk menentukan parameter optimal dan membuat model, dan set uji, yang dikecualikan dari analisis regresi dan digunakan untuk validasi. 19 , 20 Kekuatan prediksi dinilai menggunakan koefisien korelasi yang divalidasi silang ( Q 2 ) dan kesalahan prediksi akar-rata-kuadrat (RMSEP). Q 2 > 0,5 menunjukkan model yang baik, 27 dan nilai RMSEP yang lebih rendah menunjukkan kemampuan pemasangan yang lebih baik untuk rentang penuh nilai data untuk variabel dependen. 27 Berdasarkan signifikansi flavonoid pada kondisi PPFD yang berbeda yang ditunjukkan dalam hasil model PLS-DA, model PLS dibangun dengan menggunakan total kandungan flavonoid setiap sampel sebagai variabel dependen ( Y ) dan 67 metabolit, tidak termasuk flavonoid, sebagai variabel independen ( X ). Sampel bibit gandum dibagi menjadi 36 sampel pelatihan dan 18 sampel set uji (Gbr. 2A ). Model tersebut memiliki Q 2 sebesar 0,7 dan RMSEP sebesar 1,5, yang menunjukkan bahwa kadar metabolit primer diprediksi secara akurat berdasarkan total kandungan flavonoid (Gbr. 2A ). Dalam plot skor PLS, bibit gandum yang tumbuh pada PPFD 400 diprediksi mengandung kadar flavonoid tertinggi dibandingkan dengan yang tumbuh dalam kondisi lain. Metabolit signifikan dengan VIP > 1 meliputi metionina, sisteina, lutein, 13 Z – ß -karotena, ß -karotena, asam sitrat, fenilalanina, asam shikimat, asam suksinat, dan tirosin (Gbr. 2B ). Metabolit ini diklasifikasikan menjadi asam amino yang mengandung sulfur, karotenoid, zat antara siklus TCA, dan prekursor metabolisme sekunder. Intensitas cahaya memengaruhi kinerja fotosintesis, sementara asam amino yang mengandung sulfur seperti metionina dan sisteina memengaruhi produksi metabolit energi dan metabolit sekunder sebagai respons terhadap sinyal cahaya dan redoks.28 – 30 Dengan demikian, model PLS mendukung korelasi antara metabolisme primer dan peningkatan kandungan flavonoid pada bibit gandum yang dipengaruhi oleh PPFD.
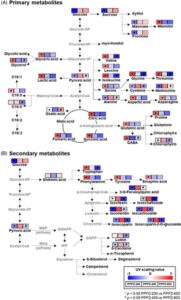
Untuk mengidentifikasi jalur metabolisme pada bibit gandum yang berubah sebagai respons terhadap PPFD, analisis pengayaan jalur dilakukan dengan membandingkan kelompok perlakuan PPFD 200 dengan 400 dan PPFD 400 dengan 800 (Gbr. 3 ). Berdasarkan metabolit yang diidentifikasi dalam basis data KEGG, enam jalur metabolisme dianggap signifikan antara bibit gandum yang tumbuh pada PPFD 200 dan 400 [−log( p ) > 7,0 dan skor dampak > 0,1]. Jalur-jalur ini meliputi siklus TCA, metabolisme glioksilat dan dikarboksilat, metabolisme piruvat, metabolisme pati dan sukrosa, metabolisme tirosin, biosintesis fenilpropanoid, dan metabolisme sistein dan metionin (Gbr. 3A ). Selain itu, antara bibit gandum yang tumbuh pada PPFD 400 dan 800, jalur yang paling terpengaruh adalah metabolisme triptofan, biosintesis karotenoid, metabolisme sistein dan metionin, serta metabolisme glisin, serin, dan treonin (Gbr. 3B ).

Lebih jauh lagi, untuk menginterpretasikan perubahan metabolik pada bibit gandum sebagai respons terhadap berbagai kondisi PPFD, representasi ringkasan jalur metabolik yang diperkaya menggunakan nilai skala UV rata-rata ditunjukkan pada Gambar 4. Intermediet siklus TCA dan sebagian besar asam amino menunjukkan kandungan yang lebih tinggi pada kondisi PPFD 200 dibandingkan dengan kondisi lain (Gambar 4A ). Keseimbangan metabolisme karbon dan nitrogen peka terhadap pertumbuhan tanaman dan pertahanan serta adaptasi terhadap faktor lingkungan. 31 , 32 Siklus TCA merupakan pusat metabolisme energi dan, bersama dengan metabolisme asam amino, paling aktif selama periode pertumbuhan awal. 32 , 33 Bibit gandum biasanya tumbuh pada PPFD 100–200 μmol m −2 s −1 , yang cocok untuk tahap vegetatif pertumbuhan daun. 32 – 34 Hasil kami menunjukkan bahwa kondisi PPFD 200 umumnya merupakan kondisi optimal untuk pertumbuhan bibit gandum, konsisten dengan temuan penelitian sebelumnya. Sebaliknya, kandungan gula (glukosa, fruktosa, manosa dan sukrosa), sisteina dan metionina meningkat pada bibit gandum PPFD 400 (Gbr. 4A ). Kandungan prekursor metabolik sekunder (asam shikimat, fenilalanina, tirosin dan triptofan), karotenoid dan flavonoid juga meningkat pada kondisi PPFD 400 (Gbr. 4B ). Fotosintesis adalah proses anabolik pada tanaman yang menghasilkan glukosa sebagai sumber energi, dan laju fotosintesis dan akumulasi gula meningkat dengan PPFD. 35 Gula adalah sumber utama metabolit sekunder; dengan demikian, akumulasi mereka mempengaruhi akumulasi metabolit sekunder, 35 yang berfungsi sebagai senyawa pertahanan tanaman terhadap stres yang disebabkan oleh berbagai faktor. 30 Selain itu, metabolisme sisteina dan metionina diketahui mengatur metabolisme sekunder tergantung pada intensitas cahaya. 36 Kandungan asam amino yang mengandung sulfur sistein dan metionina meningkat seiring dengan intensitas cahaya, dan pengendalian PPFD meningkatkan kadar sistein dan metionina, sehingga mengaktifkan biosintesis etilena, yang memediasi respons stres dalam berbagai sel. 28 , 36 , 37 Dengan kata lain, kondisi PPFD 400 menginduksi fotosintesis aktif selama pertumbuhan bibit gandum dan meningkatkan metabolisme gula dan mekanisme pertahanan, sehingga menghasilkan peningkatan kandungan flavonoid, yang merupakan senyawa antioksidan yang dikenal. Kandungan karotenoid (lutein dan ß-karoten) menurun (Gbr. 4B ), sedangkan kandungan asam oksalat meningkat (Gbr. 4A) pada bibit gandum yang tumbuh pada kondisi PPFD 800 dibandingkan dengan bibit yang tumbuh pada kondisi lain. PPFD 800 menyebabkan energi eksitasi dan fotoinhibisi yang berlebihan dibandingkan dengan kondisi pertumbuhan bibit gandum pada umumnya. 38 Karotenoid adalah pigmen fotosintetik yang kandungannya meningkat seiring dengan intensitas cahaya dan menurun pada kondisi cahaya yang berlebihan. 39 Asam oksalat merupakan indikator stres oksidatif yang disebabkan oleh cahaya yang berlebihan, 39 dan hasil kami menunjukkan bahwa perlakuan PPFD 800 mengakibatkan paparan cahaya yang berlebihan dan menghambat pertumbuhan bibit gandum. Secara keseluruhan, hasil kami menegaskan bahwa PPFD sangat memengaruhi kualitas nutrisi bibit gandum. Efek dari berbagai kondisi PPFD pada metabolit sekunder pada bibit gandum terlihat jelas, dan perubahan ini sangat berkorelasi dengan perubahan metabolit primer. Sebagai kesimpulan, kondisi PPFD 400 optimal untuk meningkatkan fotosintesis pada bibit gandum dan menghasilkan flavonoid, yang bermanfaat bagi kesehatan.
Pengaruh periode pertumbuhan terhadap kandungan metabolit bibit gandum
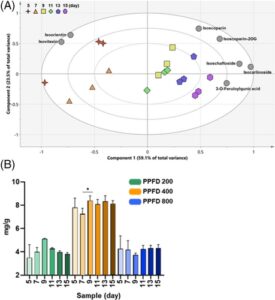
Hasil kami mengonfirmasi kualitas nutrisi flavonoid yang sangat baik pada bibit gandum yang tumbuh dalam kondisi PPFD 400. Dengan demikian, PCA dilakukan untuk menentukan efek periode pertumbuhan dalam PPFD (Gbr. 5 ). Hubungan antara sampel dan flavonoid divisualisasikan dalam biplot (Gbr. 5A ). Dalam model PCA, dua PC teratas mencakup 82,6% dari total varians (59,1% untuk PC1 dan 23,5% untuk PC2). Bibit gandum yang tumbuh selama lebih dari 9 hari dipisahkan dari yang tumbuh selama 5 dan 7 hari berdasarkan PC1. Sebagian besar flavonoid menunjukkan kandungan yang lebih tinggi pada bibit gandum yang lebih tua dari 9 hari. Lebih jauh, seperti yang ditunjukkan pada grafik batang yang mengekspresikan kandungan flavonoid total, kadar flavonoid meningkat dalam kondisi PPFD 400 dibandingkan dengan yang dalam kondisi lain (Gbr. 5B ). Kandungan flavonoid menurun pada bibit gandum yang tumbuh dalam PPFD 400 selama 7 hari dan kemudian meningkat lagi setelah 9 hari ( P < 0,05) (Gbr. 5B ). Kandungan metabolit sekunder cenderung meningkat saat tanaman dan tanaman dewasa; namun, kandungan flavonoid dalam bibit dapat bervariasi tergantung pada beberapa faktor termasuk periode pertumbuhan dan kondisi lingkungan. Pada kecambah yang dibudidayakan secara artifisial, kadar metabolit sekunder yang tinggi diproduksi dalam waktu yang relatif singkat, yaitu sekitar 6–8 hari setelah perkecambahan. 40 Perubahan ini dapat dikaitkan dengan flavonoid yang terlibat dalam berbagai aspek perkembangan tanaman, termasuk perkecambahan biji, perkembangan bibit, warna dan aroma bunga. 41 Dalam penelitian kami sebelumnya, kandungan flavonoid bibit gandum (kultivar Saekeumkang) tertinggi pada hari ke-9, diikuti oleh bibit pada hari ke-7 > hari ke-5 > hari ke-12 > hari ke-14, 17 yang konsisten dengan hasil penelitian saat ini. Dengan demikian, kondisi budidaya yang optimal untuk bibit gandum, sebagai sumber flavonoid, dipastikan adalah PPFD 400 dan pertumbuhan 9 hari.
KESIMPULAN
Kami melakukan profiling komprehensif terhadap perubahan metabolit primer dan sekunder pada bibit gandum di bawah berbagai faktor lingkungan seperti kondisi PPFD dan periode pertumbuhan. Analisis multivariat menunjukkan bahwa metabolisme bibit gandum lebih dipengaruhi oleh PPFD daripada periode pertumbuhan. Dalam model PLS-DA berdasarkan kondisi PPFD, intensitas cahaya memiliki dampak signifikan pada metabolisme flavonoid pada bibit gandum. Dalam model prediksi yang menggambarkan hubungan antara metabolit primer dan sekunder pada bibit gandum, yang tumbuh di bawah PPFD 400 diprediksi mengandung kandungan flavonoid tertinggi. Jalur metabolisme yang diperkaya dalam respons bibit gandum terhadap PPFD meliputi siklus TCA, metabolisme gula, metabolisme sistein dan metionina, biosintesis karotenoid, dan biosintesis fenilpropanoid. Zat antara siklus TCA dan sebagian besar asam amino menunjukkan kandungan yang lebih tinggi di bawah kondisi PPFD 200 daripada di bawah kondisi lain. Gula, asam piruvat, sistein, metionina, karotenoid, dan flavonoid meningkat pada bibit gandum yang dibudidayakan di bawah kondisi PPFD 400. Kandungan asam oksalat meningkat pada bibit gandum yang tumbuh dalam kondisi PPFD 800, yang menunjukkan bahwa kondisi PPFD 800 mengakibatkan paparan cahaya yang berlebihan pada bibit gandum, sehingga memengaruhi pertumbuhannya. Dibandingkan dengan bibit gandum yang tumbuh dalam kondisi lain, bibit gandum yang tumbuh pada PPFD 400 selama lebih dari 9 hari menunjukkan peningkatan fotosintesis dan kandungan flavonoid yang bermanfaat bagi kesehatan. Berdasarkan hasil ini, PPFD 200 dapat diterapkan pada periode pertumbuhan awal untuk meningkatkan perkembangan bibit, dan PPFD 400 cocok untuk periode pertumbuhan selanjutnya ketika produksi bahan fungsional seperti flavonoid perlu ditingkatkan. Hasil kami menunjukkan korelasi antara metabolit primer dan sekunder dalam kondisi PPFD pada bibit gandum. Lebih jauh, penelitian ini mendukung pengembangan strategi untuk budidaya yang dikendalikan secara lingkungan untuk meningkatkan kualitas nutrisi dan fungsional bibit gandum.



Tinggalkan Balasan