
Abstrak
Meningkatnya jumlah spesies tanaman yang terancam membutuhkan tindakan konservasi yang mendesak. Salah satu pilihannya adalah mentranslokasi tanaman kembali ke habitat aslinya. Banyak benih spesies liar disimpan di bank benih, tetapi hanya ada sedikit informasi tentang kesesuaiannya untuk translokasi tanaman. Tanaman yang tumbuh dari benih yang disimpan mungkin tidak beradaptasi dengan iklim saat ini, terutama di daerah pegunungan tempat pemanasan lebih cepat dari rata-rata global. Kami menggunakan tiga aksesi benih lama (2000–2005; “leluhur”) dan tiga aksesi benih baru-baru ini (2018–2020; “keturunan”) Viscaria alpina , spesialis pegunungan Arktik, untuk menumbuhkan tanaman yang ditranslokasi kembali ke lokasi pengumpulan benih. Kami memantau kelangsungan hidup tanaman selama 3 tahun dan mencatat sifat vegetatif, pembungaan, dan benih. Pada tahun ketiga setelah translokasi, kelangsungan hidup tinggi (96%), tetapi produksi bunga menurun. Keturunan memiliki batang berbunga lebih pendek daripada leluhur, yang menunjukkan tanaman telah menjadi lebih kecil selama 20 tahun. Jumlah bunga, massa biji, jumlah, dan perkecambahan tidak berbeda antara nenek moyang dan keturunan, yang menunjukkan bahwa ini adalah sifat yang lebih konservatif. Translokasi tanaman spesies Alpen menggunakan biji lama dan baru dikumpulkan berhasil dalam jangka pendek, yang menunjukkan bahwa biji yang disimpan selama 20 tahun dapat digunakan untuk konservasi tanaman. Namun, pemantauan jangka panjang diperlukan untuk menentukan keberhasilan translokasi ini. Tanaman keturunan lebih kecil dari nenek moyang, sehingga mereka mungkin memiliki toleransi kekeringan yang lebih baik, kemungkinan sebagai respons terhadap kondisi yang lebih kering di lokasi penelitian. Oleh karena itu, kami merekomendasikan penggunaan aksesi biji yang baru dikumpulkan untuk translokasi tanaman.
Implikasi untuk Praktik
- Tanaman yang cocok untuk dipindahkan kembali ke alam liar dapat dibudidayakan dari benih berusia 20 tahun yang disimpan di bank benih untuk proyek konservasi dan/atau restorasi, dengan tingkat kelangsungan hidup yang tinggi setelah 3 tahun.
- Pemantauan jangka panjang terhadap proyek translokasi tanaman diperlukan untuk menilai sepenuhnya hasil tindakan konservasi ini.
- Jika memungkinkan, benih yang baru dikumpulkan harus digunakan untuk translokasi konservasi karena tanaman akan lebih beradaptasi dengan kondisi iklim saat ini dan masa mendatang.
Perkenalan
Gabungan ancaman perubahan iklim, kerusakan habitat, dan fragmentasi lanskap merupakan pendorong utama hilangnya keanekaragaman hayati global, yang mengakibatkan hampir 40% spesies tumbuhan vaskular saat ini berisiko punah (Nic Lughadha et al. 2020 ). Untuk mencegah penurunan lebih lanjut dalam keanekaragaman hayati, diperlukan upaya konservasi yang cepat dan efektif. Untuk tujuan ini, Strategi Global untuk Konservasi Tumbuhan (GSPC) ditetapkan oleh Konvensi Keanekaragaman Hayati Perserikatan Bangsa-Bangsa dengan 16 target konservasi. Target 8 berfokus pada konservasi eks situ dengan tujuan menjaga minimal 75% spesies yang terancam di luar habitat alami mereka, dan 20% dari spesies ini harus tersedia untuk digunakan dalam proyek restorasi (GSPC 2012 ). Konservasi eks situ, yaitu, di luar lingkungan asli tumbuhan, dapat berupa spesimen hidup di kebun raya, kriopreservasi jaringan tumbuhan, atau penyimpanan benih di bank benih eks situ (Mounce et al. 2017 ; Walters & Pence 2021 ). Selama abad kedua puluh, koleksi hidup di kebun raya merupakan metode utama konservasi tanaman eks situ (Breman et al. 2021 ). Namun, tanaman yang tumbuh dalam kondisi non-alami diaklimatisasi ke lingkungan buatan yang lebih lembut, dan erosi genetik koleksi tanaman eks situ dapat terjadi seiring waktu, yang menyebabkan hilangnya keragaman genetik dibandingkan dengan populasi liar (Ensslin et al. 2015 ; Ensslin & Godefroid 2020 ; Wei & Jiang 2021 ). Selain itu, penggunaan materi perbanyakan eks situ hidup untuk konservasi memiliki tantangan karena tanaman yang dibudidayakan dalam kondisi antropogenik ini dapat mengalami maladaptasi ketika ditranslokasi kembali ke lingkungan alaminya karena hilangnya tekanan seleksi alam, dan oleh karena itu menunjukkan kebugaran yang berkurang (Ensslin & Godefroid 2019 ). Akibatnya, bank benih sekarang menjadi metode konservasi eks situ yang lebih disukai karena tidak memiliki risiko aklimatisasi yang terkait dengan kebun raya, dan hemat biaya, membutuhkan sedikit ruang fisik untuk menampung ribuan aksesi (Breman et al. 2021 ). Toleransi pengeringan alami benih ortodoks, yang diproduksi oleh lebih dari 90% spesies berbunga (Long et al. 2015 ), memungkinkan benih disimpan dalam jangka panjang pada suhu -18°C setelah dikeringkan pada suhu 15°C dan kelembapan relatif 15% (FAO 2014 ). Benih ortodoks dapat disimpan dalam kondisi ini selama beberapa dekade tanpa kehilangan viabilitas, membuatnya cocok untuk penyimpanan jangka panjang (Solberg et al. 2020 ). Jumlah bank benih telah meningkat pesat dalam beberapa tahun terakhir dan sekarang ada lebih dari 480 bank benih di seluruh dunia yang didedikasikan untuk konservasi spesies liar (BGCI 2023 ).
Tinjauan GSPC mengindikasikan bahwa, sementara kemajuan telah dibuat untuk mencapai target 8, pengumpulan dan penggunaan materi ex situ untuk konservasi tanaman dapat ditingkatkan (Sharrock 2020 ). Metode yang paling efektif untuk menggunakan koleksi ex situ untuk konservasi adalah dengan menanam tanaman muda atau dewasa kembali ke habitat aslinya, daripada langsung menabur benih (Dalrymple et al. 2012 ; Liu et al. 2015 ; Silcock et al. 2019 ). Translokasi didefinisikan sebagai pergerakan yang disengaja, untuk tujuan konservasi, individu dari satu lokasi ke lokasi lain untuk memperkuat atau memperkenalkan kembali populasi dalam jangkauan aslinya atau memperkenalkan spesies di luar jangkauan alaminya dengan kolonisasi berbantuan atau penggantian ekologis (World Conservation Union 2013 ). Dalam literatur ilmiah, ada beberapa keberhasilan translokasi tanaman yang menyelamatkan spesies yang terancam (misalnya Fenu et al. 2019 ; Monks et al. 2019 ; Hyvärinen 2020 ), meskipun informasi tentang translokasi tanaman menggunakan benih yang disimpan secara ex situ masih terbatas (White et al. 2023 ). Efektivitas tindakan translokasi yang sebenarnya sulit ditentukan karena hasil positif sering kali dilaporkan secara berlebihan dibandingkan dengan kegagalan (Godefroid et al. 2011 ). Selain itu, kuantifikasi hasil translokasi dibatasi oleh kurangnya pemantauan jangka panjang dan tujuan yang tidak ditentukan, yang mengarah pada pandangan yang terlalu optimis terhadap hasil translokasi tanaman (Godefroid et al. 2011 ). Ada banyak tantangan dan risiko yang terkait dengan translokasi tanaman kembali ke lingkungan alaminya, termasuk ketersediaan habitat yang sesuai, kondisi pertumbuhan yang tidak menguntungkan yang disebabkan oleh perubahan iklim, dan sumber bahan yang sesuai untuk translokasi (Ensslin et al. 2015 ; Godefroid et al. 2016 ; Commander et al. 2018 ). Faktor terakhir ini merupakan elemen penting dalam merencanakan tindakan translokasi dan sangat penting untuk keberhasilan hasilnya (Godefroid et al. 2016 ). Mengumpulkan bahan tanaman segar dari alam liar dapat menjadi sulit karena faktor-faktor seperti set benih yang rendah dan/atau kualitas, dan pembatasan pengambilan sampel untuk melindungi populasi alami dari panen berlebihan (Pedrini et al. 2020 ). Akibatnya, koleksi ex situ adalah sumber alternatif yang berguna, terutama untuk spesies yang terancam (Abeli et al. 2020 ).
Karena tantangan yang terkait dengan penggunaan bahan dari koleksi hidup, benih yang disimpan di bank benih ex situ merupakan bahan sumber yang berguna untuk menumbuhkan tanaman untuk ditranslokasi ke habitat alaminya. Koleksi ini memiliki manfaat karena tidak terpapar pada kondisi buatan kebun botani dan jika viabilitas benih tinggi dan pengambilan sampel benih mewakili keseluruhan populasi, keragaman genetik koleksi tersebut tetap terjaga. Namun, tanaman yang tumbuh dari benih yang disimpan selama bertahun-tahun dalam kondisi bank benih mungkin tidak lagi beradaptasi dengan iklim saat ini ketika dipindahkan kembali ke habitat alami (Schoen & Brown 2001 ; Ensslin et al. 2015 ; Karitter et al. 2024 ). Selain itu, jika benih kehilangan viabilitas seiring waktu dalam penyimpanan dan jika penurunan ini memiliki dasar genetik, apa yang disebut “fraksi tak terlihat” mungkin ada, yang berpotensi menyebabkan hilangnya variasi genetik (Weis 2018 ). Studi terbaru yang membandingkan tanaman dari populasi liar dan tanaman dari koleksi eks situ telah menunjukkan bahwa tanaman dari populasi liar telah beradaptasi dari waktu ke waktu sebagai respons terhadap kondisi iklim saat ini (Rauschkolb et al. 2022 ; Ensslin et al. 2023 ). Ini menyiratkan bahwa tanaman dari sumber eks situ mungkin tidak cocok untuk ditranslokasi kembali ke alam liar. Risiko ini mungkin sangat tinggi di daerah yang mengalami perubahan iklim yang cepat, di mana seleksi alam dapat bertindak lebih cepat. Namun, untuk spesies langka atau terancam, bahan eks situ, terutama benih, mungkin merupakan satu-satunya sumber yang tersedia, jadi penting untuk memahami bagaimana tanaman yang tumbuh dari bahan ini akan tumbuh ketika ditranslokasi kembali ke habitat alami.
Tujuan dari penelitian ini adalah untuk mentranslokasi tanaman spesialis Arktik-Alpin Viscaria alpina , yang telah dibudidayakan dari benih yang disimpan dalam kondisi bank benih untuk jangka waktu yang berbeda, kembali ke lokasi pengumpulan benih dan memantau keberhasilan translokasi. Keuntungan mengembalikan tanaman kembali ke lokasi pengumpulan benih asli adalah bahwa setiap perbedaan yang diamati dapat dikaitkan dengan perubahan yang telah terjadi dari waktu ke waktu daripada karena lokasi yang berbeda. Dengan pemanasan iklim yang terjadi lebih cepat di daerah Alpen dibandingkan dengan rata-rata global (Hock et al. 2019 ), tanaman yang diperbanyak dari benih ex situ yang telah disimpan selama bertahun-tahun dan dipindahkan kembali ke lingkungan Alpen mungkin kurang cocok dengan kondisi yang lebih hangat dan lebih kering. Spesies Alpen dianggap sangat berisiko dengan pemanasan iklim karena mereka beradaptasi dengan lingkungan yang dingin dan keras dan sering kali membutuhkan lapisan salju dan suhu rendah untuk menyelesaikan siklus hidupnya (Körner 2003 ). Survei terkini terhadap bank benih di seluruh dunia menunjukkan bahwa meskipun bank secara aktif melakukan translokasi tanaman menggunakan koleksi mereka, hasil translokasi tidak dipublikasikan secara rutin (White et al. 2023 ). Oleh karena itu, hanya ada sedikit informasi yang tersedia tentang kegunaan benih yang disimpan di bank benih untuk translokasi tanaman guna mendukung konservasi. Dengan mentranslokasi tanaman spesies pegunungan yang tumbuh dari aksesi benih yang disimpan secara eks situ yang berbeda dan menyelidiki kelangsungan hidup dan hasil reproduksi, kami akan berkontribusi pada pemahaman yang lebih baik tentang kegunaan benih eks situ untuk translokasi tanaman di daerah pegunungan tinggi.
Metode
Studi Spesies dan Lokasi Pengumpulan/Translokasi Benih
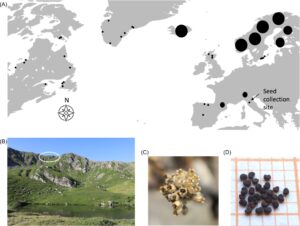
Viscaria alpina (L.) G. Don (Caryophyllaceae) adalah spesies tanaman tahunan alpen-arktik yang terbentuk dari semi-roset daun dengan batang berbunga (Nagy 2013 ). Tanaman ini sangat toleran terhadap stres, tumbuh terutama di tanah berbatu, dengan distribusi di belahan bumi utara di seluruh Eropa, Greenland, dan Amerika Utara di lokasi alpen dan subarktik (Gbr. 1A ; Nagy 2013 ). Spesies ini umumnya hidup selama 3–10 tahun dan bersifat iteroparous, berbunga dalam beberapa tahun (Nagy & Proctor 1996 ). Meskipun V. alpina memiliki distribusi yang relatif besar, di Italia tanaman ini diklasifikasikan sebagai rentan menurut kriteria World Conservation Union (Abeli et al. 2009 ). Benih V. alpina telah dikumpulkan sejak tahun 2000 selama beberapa tahun pada saat penyebaran alami (Agustus/September) dari dekat puncak Monte Prado di Apennini utara, Italia (44°14′N 10°24′E, 2054 m dpl; Gbr. 1B & 1C ). Lokasi pengumpulan benih terletak di Taman Nasional Appennino Tosco-Emiliano, yang berada di batas wilayah selatan untuk spesies ini. Populasi V. alpina di Monte Prado dianggap sebagai peninggalan glasial, dan meskipun populasinya kecil, terisolasi, dan terletak di area terbatas, populasinya tetap stabil selama pemantauan jangka panjang (Abeli et al. 2009 , 2012 ). Pada setiap pengambilan sampel, ribuan benih dikumpulkan dari minimal 30 individu dan disimpan dalam kondisi bank benih standar FAO: −18°C setelah dikeringkan pada suhu 15°C dan kelembaban relatif 15% (FAO 2014 ) di Bank Benih Universitas Pavia, Pavia, Italia (Gbr. 1D ). Benih yang dikumpulkan antara tahun 2000 dan 2005 telah disimpan di Bank Benih Milenium, Royal Botanic Gardens Kew, Inggris Raya, sebelum dipindahkan ke Universitas Pavia pada tahun 2019 dalam paket aluminium tertutup dan ditempatkan di freezer bank benih.
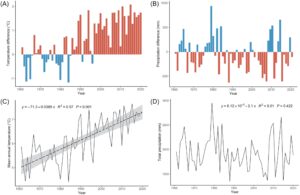
Iklim di lokasi pengumpulan benih diekstrapolasi dari basis data daring dengan data suhu dan curah hujan yang berasal dari tahun 1961 (Antolini et al. 2016 ; https://arpaeprv.datamb.it/dataset/erg5-eraclito ). Membandingkan iklim periode sebelum pemanasan (1961–1990; WMO 2015 ) dengan iklim selama periode pengumpulan benih (2000–2020) menunjukkan bahwa semua tahun lebih hangat daripada rata-rata dan 13 dari 20 tahun terakhir memiliki curah hujan yang lebih rendah daripada rata-rata (Gbr. 2A & 2B ). Selama periode ini suhu telah menghangat sebesar 0,6°C, yang setara dengan 0,3°C per dekade, lebih cepat daripada rata-rata global sebesar 0,2°C per dekade (Allen et al. 2018 ) sementara curah hujan bervariasi setiap tahun (Gbr. 2C & 2D ).
Persiapan Bibit
Pada bulan Juni 2021, tanaman untuk translokasi dibudidayakan dari benih enam aksesi yang berbeda, dengan yang dari tahun 2000, 2001, dan 2005 digolongkan sebagai “nenek moyang” dan dari tahun 2018, 2019, dan 2020 sebagai “keturunan.” Untuk setiap aksesi, 50 benih ditabur pada agar air suling 1% pada cawan Petri 90 mm. Benih-benih ini ditempatkan dalam inkubator pada suhu 25°C dengan periode cahaya terang/gelap 12 jam, yang sebelumnya terbukti menjadi kondisi perkecambahan optimal untuk V. alpina (Mondoni et al. 2018 ). Perkecambahan awal tinggi lebih dari 95% untuk semua aksesi. Ketika benih telah berkecambah (didefinisikan sebagai radikula ≥2 mm panjang) mereka dipindahkan dengan hati-hati ke pot (4,5 × 4 × 11 cm) yang berisi tanah pot standar. Nampan perbanyakan disimpan di bawah lampu dengan radiasi aktif fotosintesis sebesar 300 μmol m −2 s −1 (model Ambralight AE80, Italia), yang diatur pada periode cahaya terang/gelap selama 12 jam. Suhu tetap konstan pada 23°C dan tanah dijaga tetap lembap. Aksesi didistribusikan secara acak di seluruh nampan perbanyakan dan nampan dirotasi setiap 3–4 hari untuk memastikan distribusi cahaya yang merata.
Translokasi Tanaman
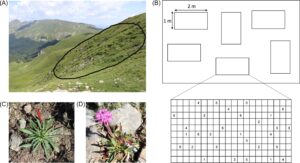
Bahasa Indonesia: Ketika tanaman berusia sekitar 7 minggu, mereka dipindahkan ke Viote Alpine Botanical Garden, Italia (46°02′N 11°03′E, 1540 m dpl) untuk mengaklimatisasi mereka dengan lingkungan yang lebih mirip dengan yang mereka alami di alam. Tanaman ditranslokasi ke lokasi pengumpulan benih di Monte Prado (detail di atas) pada akhir musim tanam untuk meminimalkan kehilangan tanaman yang dapat terjadi pada bulan-bulan yang lebih kering dan panas di bulan Juli dan Agustus. Translokasi dilakukan dengan menggunakan plot ulangan yang terdiri dari enam plot 2 × 1 m 2 dengan arah lereng yang sama, kemiringan, dan tutupan vegetasi yang sama (Gbr. 3A ). Setiap plot berisi lima tanaman per aksesi, jadi 30 individu per plot (Gbr. 3B ). Tanaman ditempatkan secara acak dalam setiap plot dan ditandai dengan pengenal yang sesuai dengan aksesi benih (Gbr. 3C & 3D ). Plot dipilih di area tempat V. alpina tumbuh liar, sehingga kondisinya cocok untuk pertumbuhan. Tanaman dipindahkan pada tanggal 06 September 2021 saat berusia sekitar 11 minggu. Pencatat potensial air dan suhu (MicroLog SP3, Environmental Measurement Systems, Brno, Republik Ceko) ditempatkan di empat plot untuk memantau kondisi tanah selama percobaan.
Pengukuran Sifat
Kelangsungan hidup tanaman setelah musim dingin dicatat dan sifat reproduksi diukur selama musim berbunga (Juli) selama 3 tahun setelah translokasi, 2022–2024. Untuk setiap individu, status pembungaan dicatat bersama dengan tinggi setiap batang berbunga dan jumlah bunga. Setelah musim berbunga (akhir Agustus/awal September 2022–2024) kelangsungan hidup tanaman dicatat. Setelah tahun pertama, benih dikumpulkan dari individu dengan buah matang. Massa benih untuk semua individu yang dikumpulkan ditentukan dengan mengukur massa 250 benih dalam lima replikasi 50 benih dan menghitung rata-ratanya. Benih yang tersisa ditimbang dan ditambahkan ke massa total. Jumlah benih untuk setiap individu yang dikumpulkan setelah tahun pertama dihitung dari massa jumlah benih yang diketahui, massa benih total sampel, dan berat rata-rata ditambah nilai yang dihitung dari batas kepercayaan 95% pada kesalahan standar. Metode ini mempertimbangkan potensi kesalahan pengambilan sampel. Untuk memeriksa perkecambahan, enam ulangan per aksesi, dipilih secara acak dari plot yang berbeda, disemai dalam rangkap tiga (3 × 20 benih) pada agar air suling 1% dan ditempatkan pada suhu 25°C dengan periode foto terang/gelap 12 jam. Benih diperiksa untuk perkecambahan (panjang radikula ≥2 mm) secara teratur, dan setelah 30 hari, benih yang belum berkecambah dipotong terbuka untuk menentukan apakah benih tersebut layak. Benih yang mati atau kosong dikeluarkan dari analisis. Waktu perkecambahan rata-rata adalah ukuran kecepatan perkecambahan dan dihitung menggunakan metode dalam Lozano-Isla et al. ( 2019 ). Total perkecambahan dan waktu perkecambahan rata-rata dapat memberikan indikasi dinamika perkecambahan.
Analisis Statistik
Model campuran linear untuk setiap sifat (tinggi batang berbunga, jumlah bunga, massa biji, dan jumlah) dilakukan. Sifat tersebut adalah variabel respons, sedangkan efek asal temporal biji (yaitu nenek moyang atau keturunan) dan tahun setelah translokasi dan interaksinya adalah efek tetap. Biji hanya dikumpulkan setelah tahun pertama, jadi untuk massa biji dan jumlah biji, tahun setelah translokasi tidak termasuk dalam model. Untuk memperhitungkan pengaturan eksperimen, plot dimasukkan sebagai efek acak. Uji Tukey post hoc dilakukan untuk perbandingan berpasangan asal temporal biji serta untuk tahun setelah translokasi, menggunakan paket emmeans (Lenth 2022 ) dan multcomp (Hothorn et al. 2008 ). Perkecambahan dianalisis dengan model linear umum dengan fungsi tautan logit kuasi-binomial, termasuk perkecambahan akhir sebagai variabel respons dan asal temporal biji sebagai variabel penjelas. Kelangsungan hidup dan pembungaan tanaman dianalisis menggunakan model campuran linier umum dengan fungsi tautan logit binomial menggunakan estimasi kemungkinan maksimum dengan perkiraan Laplace untuk distribusi data non-normal. Kelangsungan hidup atau pembungaan tanaman adalah variabel respons, asal temporal benih dan tahun setelah translokasi adalah variabel penjelas, termasuk interaksinya. Plot dimasukkan sebagai efek acak. Semua analisis statistik dilakukan dalam R versi 4.2.2 (Tim Inti R 2022 ): model dilakukan dalam paket lme4 (Bates et al. 2015 ); asumsi diperiksa menggunakan paket DHARMa (Hartig & Lohse 2022 ); data perkecambahan dianalisis dengan paket GerminaR (Lozano-Isla et al. 2019 ) dan grafik digambar dalam ggplot2 (Wickham 2016 ) dengan palet yang sesuai untuk buta warna dari RColorBrewer (Neuwirth 2022 ). Semua asumsi model terpenuhi.
Hasil
Kelangsungan hidup tanaman 3 tahun setelah translokasi tinggi, dengan 96% tanaman masih hidup setelah musim panas ketiga. Akan tetapi, terdapat perbedaan signifikan dalam jumlah individu yang berbunga antar tahun, dengan 98% pembungaan pada tahun pertama menurun menjadi 87% pada tahun kedua dan kemudian meningkat menjadi 92% pembungaan pada tahun ketiga, meskipun tidak ditemukan perbedaan antara nenek moyang dan keturunan (Tabel S1 ). Perkecambahan benih yang dikumpulkan setelah pembungaan pertama tidak berbeda antara nenek moyang dan keturunan dalam perkecambahan total atau waktu perkecambahan rata-rata (Tabel S1 ).
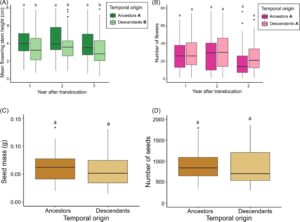
Terdapat bukti bahwa keturunan memiliki batang berbunga yang jauh lebih pendek daripada nenek moyang, dan perbedaan ini tetap ada setelah tahun ketiga pemantauan (Tabel S2 & S3 ; Gambar 4A ). Selain itu, tanaman memiliki lebih sedikit bunga pada tahun ketiga dibandingkan dengan tahun-tahun sebelumnya, tetapi tidak ada perbedaan antara nenek moyang dan keturunan (Tabel S2 & S3 ; Gambar 4B ). Tidak ada bukti perbedaan antara leluhur dan keturunan dalam jumlah dan massa benih yang dihasilkan setelah tahun pertama (Tabel S4 ; Gambar 4C & 4D ).
Diskusi
Dalam penelitian ini, kami menyelidiki hasil translokasi tanaman menggunakan benih yang disimpan di bank benih dari aksesi yang lebih tua dan yang baru dikumpulkan sebagai bahan sumber. Tingkat kelangsungan hidup tanaman yang tinggi setelah translokasi kembali ke lokasi pengumpulan benih menunjukkan bahwa benih yang disimpan di bank benih cocok untuk konservasi spesies spesialis Arktik/Alpin Viscaria alpina , dan mungkin spesies Alpen lainnya. Namun, tinggi batang berbunga yang lebih pendek pada keturunan dibandingkan dengan nenek moyang menunjukkan bahwa spesies ini telah berevolusi menjadi lebih kecil dari waktu ke waktu.
Dalam hal kelangsungan hidup tanaman, translokasi V. alpina kembali ke lokasi pengumpulan benih berhasil. Kelangsungan hidup tinggi, dengan 96% tanaman masih hidup setelah tahun ketiga pemantauan dan tidak ada perbedaan yang diamati antara nenek moyang dan keturunannya. Penelitian sebelumnya yang menyelidiki kesesuaian benih yang disimpan untuk translokasi tanaman menunjukkan bahwa tanaman dari sumber eks situ yang lebih tua kurang beradaptasi dengan iklim saat ini dan oleh karena itu mungkin tidak cocok untuk ditranslokasi kembali ke alam liar (Ensslin et al. 2015 ; White et al. 2023 ). Sementara kelangsungan hidup tanaman 3 tahun setelah translokasi tinggi, jumlah tanaman yang berbunga menurun setelah tahun pertama, dan jumlah rata-rata bunga per tanaman lebih rendah pada tahun ketiga. Meskipun persentase tanaman yang masih menghasilkan bunga tinggi (lebih dari 85%), penurunan yang diamati mungkin menunjukkan bahwa, dalam jangka panjang, translokasi ini tidak akan berhasil. Namun, fluktuasi jumlah batang berbunga pada populasi alami V. alpina di lokasi penelitian biasanya tinggi antar tahun (Abeli et al. 2012 ).
Tinjauan sebelumnya tentang hasil translokasi tanaman telah menunjukkan bahwa kelangsungan hidup dan hasil reproduksi menurun seiring waktu, terutama ketika pemantauan jangka panjang lebih dari 10 tahun dilakukan (Dalrymple et al. 2011 ; Godefroid et al. 2011 ). Salah satu alasan penurunan pembungaan yang kami amati dapat dikaitkan dengan kondisi iklim pada musim semi dan panas tahun 2022. Musim ini adalah yang terkering yang pernah tercatat di sebagian besar wilayah Eropa, dengan kekeringan parah yang diperburuk oleh pemanasan iklim antropogenik yang memengaruhi banyak wilayah, termasuk lokasi penelitian kami di Italia utara (Bonaldo et al. 2022 ; Faranda et al. 2023 ). Sebagai respons terhadap tekanan kekeringan tahun sebelumnya, bisa jadi lebih sedikit tanaman yang berbunga; dan meskipun persentase pembungaan lebih tinggi pada tahun ketiga, hasil reproduksi (jumlah bunga) lebih rendah. Fluktuasi tahunan yang sebelumnya dilaporkan dalam jumlah batang berbunga pada populasi V. alpina alami di lokasi studi kemungkinan terkait dengan durasi tutupan salju, sehingga secara tidak langsung terkait dengan tingkat kekeringan di awal musim panas (data Abeli yang tidak dipublikasikan). Respons serupa terhadap kekeringan telah diamati pada spesies alpine di mana curah hujan yang lebih rendah pada tahun sebelumnya mengurangi pembungaan pada tahun berikutnya (Dolezal et al. 2020 ). Selain itu, spesies alpine di dataran tinggi merespons kekeringan lebih kuat daripada yang berada di dataran rendah, dengan laju pertumbuhan yang lebih lambat dan hasil reproduksi yang lebih rendah (Rosbakh et al. 2017 ). Karena V. alpina adalah spesies menahun, mungkin saja tanaman menghemat sumber daya dengan tidak berbunga setelah mengalami stres, sehingga meningkatkan peluang bertahan hidup mereka. Memang, kelangsungan hidup tanaman tidak menurun, yang menunjukkan bahwa spesies ini akan mampu mengatasi peningkatan kekeringan yang diperkirakan akan terjadi dengan pemanasan iklim di pegunungan Mediterania (Nogués-Bravo et al. 2008 ). Namun, karena jumlah tanaman yang berbunga dan jumlah bunga per tanaman menurun, mungkin sebagai respons terhadap kekeringan, hal ini dapat memengaruhi hasil reproduksi dan perekrutan tanaman di masa mendatang. Pemantauan jangka panjang akan diperlukan untuk menilai sepenuhnya hasil translokasi ini.
Tanaman Viscaria alpina yang tumbuh dari benih yang baru dikumpulkan memiliki batang berbunga yang lebih pendek daripada yang berasal dari benih yang lebih tua, dan perbedaan ini bertahan hingga musim berbunga ketiga. Akar tunggang yang kuat di V. alpina memungkinkan kelangsungan hidup selama musim dingin, dan selama periode ini, sebagian besar dedaunan mati, dengan daun baru dan batang berbunga muncul di musim semi (Nagy 2013 ). Karena keturunan mempertahankan batang berbunga yang lebih pendek daripada leluhur di musim tanam ketiga, tampaknya sifat ini tidak berubah setiap tahun tetapi telah berevolusi dalam populasi liar selama 20 tahun. Tinggi tanaman dan biomassa berkorelasi positif (Harrison & LaForgia 2019 ) sehingga batang berbunga yang lebih pendek menyiratkan bahwa tanaman lebih kecil dan memiliki biomassa di atas tanah yang lebih rendah. Sayangnya, tidak mungkin untuk mengukur biomassa di lapangan karena ini akan melibatkan pengumpulan seluruh tanaman. Namun, penelitian sebelumnya menggunakan tanaman dari aksesi benih V. alpina yang sama yang tumbuh dalam kondisi laboratorium menemukan bahwa keturunan memiliki biomassa di atas tanah yang lebih rendah daripada leluhur, bahkan di bawah ketersediaan sumber daya penuh (White et al. 2023 ). Bukti ini, bersama dengan batang bunga yang lebih pendek yang diamati di sini, menunjukkan bahwa populasi V. alpina ini ini telah menjadi lebih kecil selama 20 tahun terakhir. Alasan mengapa tanaman dari aksesi benih yang lebih baru menjadi lebih kecil bisa jadi merupakan adaptasi terhadap peningkatan kekeringan yang dialami di lokasi penelitian. Lokasi ini memiliki suhu yang lebih tinggi dari rata-rata dan curah hujan yang lebih rendah selama periode pengumpulan benih. Kedua faktor ini berkontribusi terhadap tanaman dari beberapa tahun terakhir yang terpapar pada peningkatan frekuensi dan tingkat keparahan kekeringan. Sebagai tanggapan, tampaknya tanaman telah menjadi lebih kecil untuk meningkatkan kemungkinan bertahan hidup dalam ketersediaan air yang rendah. Tanaman yang lebih kecil mengatasi kekeringan dengan lebih baik karena memiliki evapotranspirasi air yang lebih rendah dari daunnya (Lloret et al. 1999 ; Markesteijn & Poorter 2009 ). Hal ini selanjutnya didukung oleh Rauschkolb et al. ( 2022 ) yang menemukan kelangsungan hidup yang lebih tinggi dan pengurangan ukuran tanaman dalam kekeringan pada keturunan dibandingkan dengan nenek moyang ketika menyelidiki perbedaan antara tanaman yang tumbuh dari benih yang disimpan di luar situ berusia minimal 21 tahun dan dari benih yang baru dikumpulkan.
Selain sifat di atas tanah, mungkin saja tanaman turunannya lebih kecil karena mereka lebih banyak berinvestasi pada biomassa di bawah tanah. Ini adalah strategi umum di bawah kekeringan di mana tanaman telah terbukti mengembangkan akar yang lebih dalam untuk meningkatkan penyerapan air dari tanah (Pérez-Ramos et al. 2013 ; Harrison & LaForgia 2019 ), meskipun respons ini tidak universal (Larson et al. 2020 ). Sayangnya, kami tidak dapat mengukur sifat-sifat akar karena kami ingin terus memantau tanaman yang ditranslokasi selama beberapa tahun ke depan. Jumlah bunga tidak berbeda antara nenek moyang dan keturunan, yang menunjukkan bahwa sifat ini lebih konservatif daripada ukuran tanaman. Selain itu, benih yang dikumpulkan setelah tahun pertama tidak berubah dalam jumlah, massa, total perkecambahan, atau waktu perkecambahan rata-rata antara nenek moyang dan keturunan. Pekerjaan sebelumnya pada spesies ini telah menunjukkan bahwa benih dari keturunan lebih berat daripada nenek moyang (White et al. 2023 ); Namun, temuan ini berasal dari tanaman yang dibudidayakan dalam kondisi buatan yang mungkin tidak mencerminkan kompleksitas yang terjadi di alam (Kreyling & Beier 2013 ).
Saat mengolah tanaman yang tumbuh dari benih yang disimpan secara eks situ, penggunaan generasi penyegaran direkomendasikan untuk meminimalkan efek maternal dan waktu penyimpanan (Franks et al. 2019 ). Kami tidak menggunakan generasi penyegaran karena V. alpina adalah spesies menahun yang tidak berbunga pada tahun pertama sehingga batasan waktunya akan terlalu besar. Penelitian sebelumnya pada spesies menahun juga tidak menyertakan generasi penyegaran karena alasan ini (misalnya Everingham et al. 2021 ; Rauschkolb et al. 2023 ). Selain itu, tanaman harus tumbuh dalam kondisi budidaya yang berarti mereka mungkin dapat menyesuaikan diri dengan lingkungan non-alami (Ensslin et al. 2015) ; Ensslin & Godefroid 2020 ) dan oleh karena itu tidak mewakili tanaman yang dihasilkan dari benih yang disimpan. Daya berkecambah awal V. alpina yang tinggi yang kami amati (>95%) juga menunjukkan bahwa sangat sedikit benih yang kehilangan viabilitas selama penyimpanan, oleh karena itu keberadaan apa yang disebut “fraksi tak terlihat” terbatas (Weis 2018 ). Akibatnya, kami yakin bahwa tidak ada hilangnya variabilitas genetik akibat kerusakan selama penyimpanan.
Tingkat kelangsungan hidup V. alpina yang tinggi setelah translokasi menjanjikan untuk konservasi bank benih eks situ untuk spesies ini, dan mungkin spesies lain, spesialis pegunungan Alpen. Tampaknya V. alpina sangat toleran terhadap stres dan tanaman yang tumbuh dari benih berusia minimal 20 tahun dapat ditranslokasi kembali ke alam liar. Sementara hasil reproduksi menurun selama periode pemantauan, fluktuasi alami umum terjadi pada populasi liar (Abeli et al. 2012 ). Oleh karena itu, pemantauan jangka panjang diperlukan untuk mengevaluasi keberhasilan translokasi ini secara menyeluruh. Keturunan tampaknya lebih beradaptasi terhadap kekeringan daripada nenek moyang, suatu keuntungan di bawah suhu yang lebih tinggi dan peningkatan frekuensi kekeringan yang dialami lokasi penelitian dan yang kemungkinan akan terus dialami oleh wilayah pegunungan Alpen di masa mendatang (Gobiet et al. 2014 ; Baronetti et al. 2022) ; Kotlarski et al. 2023 ). Oleh karena itu, akan bermanfaat untuk menggunakan tanaman yang tumbuh dari aksesi benih yang dikumpulkan terakhir untuk ditranslokasi kembali ke alam. Sayangnya, hal ini tidak selalu memungkinkan, khususnya untuk spesies yang terancam punah, yang mungkin memiliki ketersediaan benih terbatas dan risiko panen berlebihan terhadap populasi lokal tinggi (Liu et al. 2020 ). Namun, kami telah menunjukkan bahwa tanaman yang tumbuh dari benih yang lebih tua dapat tumbuh, dengan tingkat kelangsungan hidup yang tinggi setelah translokasi, tetapi mungkin kurang cocok untuk iklim yang lebih hangat dan kering di masa mendatang. Kami menyarankan, jika memungkinkan, agar bank benih meningkatkan frekuensi pengumpulannya untuk memastikan bahan yang paling sesuai tersedia untuk translokasi.



Tinggalkan Balasan