
Abstrak
Mengidentifikasi daerah genomik dalam seleksi sangat penting untuk memahami sejarah evolusi ayam domestik. Ayam asli Semenanjung Arab (AP) sebagian besar ditemukan di luar ruangan, dipelihara bersama ternak lain untuk tujuan produksi. Burung-burung ini menunjukkan ketahanan tinggi terhadap suhu ekstrem (panas dan dingin), khas lingkungan gurun. Tekanan seleksi yang bertanggung jawab atas adaptasi lokal yang unik pada burung-burung ini sebagian besar masih belum teridentifikasi. Di sini, kami bertujuan untuk menyelidiki keragaman genom dan struktur 15 populasi ayam asli termasuk 13 populasi dari AP ( n = 5), Ethiopia ( n = 6), dan Republik Rakyat Tiongkok ( n = 2). Kami juga menyertakan dua populasi ayam komersial, Fayoumi (dipilih untuk toleransi panas) dan Chantecler (dikenal karena toleransi dinginnya). Analisis komponen utama (PC) memisahkan semua populasi berdasarkan daerah asal geografisnya. PC1 memisahkan populasi Ethiopia dari populasi Tiongkok dan AP, sementara PC2 memisahkan populasi AP dari Chantecler, dan populasi Ethiopia dari Dulong dan Chantecler. Tanda-tanda genom secara luas dari analisis mengidentifikasi banyak kandidat daerah di bawah seleksi positif. Mereka termasuk gen yang mungkin terkait dengan termotoleransi. Ini terlibat dalam keseimbangan energi dan metabolisme ( SUGCT , HECW1 , MMADHC ), apoptosis sel ( APP , SRBD1 , NTN1 , PUF60 , SLC26A8 , DAP , SUGCT ), angiogenesis ( RYR2 , LDB2 , SOX5 ), perlindungan kulit terhadap radiasi matahari ( FZD10 , BCO2 , WNT5B , COL6A2 , SIRT1 ) serta pertumbuhan ( NELL1 ). Temuan kami menunjukkan bahwa populasi ayam Arab memiliki polimorfisme kumpulan gen yang berbeda dalam kaitannya dengan adaptasi mereka terhadap lingkungan iklim yang keras di AP.
PERKENALAN
Populasi ayam asli, yang terbentuk selama ribuan tahun melalui adaptasi, menawarkan wawasan menarik tentang tanda-tanda genomik seleksi alam dalam menanggapi lingkungan termal yang bervariasi. Burung-burung yang mudah beradaptasi ini telah mengembangkan sifat-sifat unik yang memungkinkan mereka untuk bertahan hidup dan bereproduksi dalam iklim yang beragam, menyoroti interaksi rumit antara evolusi genetik dan tekanan lingkungan (Lawler, 2016 ). Misalnya, Perini et al. ( 2020 ) meninjau jalur molekuler yang terkait dengan stres panas, sementara Juiputta et al. ( 2023 ) membahas strategi genetik untuk meningkatkan toleransi panas dan mempertahankan produktivitas dalam peternakan unggas tropis. Selain itu, Nawaz et al. ( 2024 ) menilai gen dan jalur penting yang terkait dengan toleransi panas, seperti respons syok panas, sistem pertahanan antioksidan, fungsi imun, dan homeostasis seluler.
Mereka tumbuh subur di iklim panas, seperti daerah kering di sub-Sahara Afrika atau gurun terik di Jazirah Arab (AP), setelah mengembangkan serangkaian adaptasi untuk menahan tekanan panas yang tiada henti (Fathi, Al-Homidan, Abou-Emera & Al-Moshawah, 2017 ; Kanyama et al., 2022 ). Adaptasi ini mencakup berbagai polimorfisme fisiologis, perilaku, dan genetik yang ditujukan untuk mempertahankan homeostasis di lingkungan bersuhu tinggi (Cheng & Muir, 2005 ). Yang menonjol di antaranya adalah mekanisme termoregulasi yang efisien, seperti peningkatan reflektansi bulu dan kapasitas untuk menghilangkan panas melalui perilaku seperti mencari tempat teduh dan terengah-engah (Soleimani et al., 2011 ).
Studi genomik telah mengungkap tanda tangan genetik yang terkait dengan toleransi panas, menekankan variasi substansial dalam gen yang bertanggung jawab untuk beradaptasi dengan stres panas di antara populasi ayam asli. Misalnya, Guo et al. ( 2022 ) mengidentifikasi beberapa sapuan selektif yang mengandung gen dalam seleksi ( FABP2 , RAMP3 , SUGCT , dan TSHR ) pada ayam asli dari daerah tropis (Tiongkok utara, Indonesia, Sri Lanka, dan Thailand), yang mungkin terkait dengan adaptasi terhadap suhu sekitar yang lebih tinggi. Mereka secara khusus menemukan mutasi salah arti pada TSHR yang dapat meningkatkan toleransi panas pada ayam. Zhuang et al. ( 2020 ) mengidentifikasi gen yang terkait dengan termotolerans pada ayam asli Taiwan, termasuk gen yang terlibat dalam perbaikan DNA, respons stres seluler, apoptosis, dan stres oksidatif metabolik. Lawal et al. ( 2018 ) mengamati bahwa gen KCNMA1 berada dalam wilayah tanda tangan seleksi pada populasi ayam Arab Saudi dan Sri Lanka. Gen ini terkait dengan respons terhadap tantangan hipoksia dan regulasi kontraksi otot polos. Beberapa gen yang terlibat dalam adaptasi terhadap iklim juga diidentifikasi pada ayam kampung Tiongkok, termasuk APP , FABP1 , SMYD1 , UBE4B , NALCN , PDGFRA , NRP1 , CORIN , CLPTM 1L , CRADD , PARK2 , SIM2 , AHR , ESRRG , IL18 , dan BVES (Gu et al., 2020 ). Pada ayam kampung Nigeria, gen yang mungkin terkait dengan adaptasi terhadap stres panas (misalnya, ILF3 , HSF1 , SLC44A2 , SFTPB , HIF3A , CDC37 , dan TSHR ) baru-baru ini dilaporkan (Rachman et al., 2024 ).
Selain itu, investigasi genomik telah mengidentifikasi varian genetik yang terkait dengan adaptasi dingin, termasuk varian yang mengatur perkembangan bulu, metabolisme lipid, dan termogenesis yang disebabkan oleh dingin (Wang et al., 2015 ). Adaptasi ini sangat diperlukan untuk bertahan hidup di iklim dingin, di mana konservasi energi dan retensi panas sangat penting. Dalam analisis genom yang komprehensif oleh Xu et al. ( 2021 ), dua kandidat daerah terpilih mencakup gen yang mungkin terkait dengan adaptasi terhadap stres dingin di Chantecler telah diidentifikasi. Daerah ini meliputi gen ME3 , yang terlibat dalam metabolisme lemak, dan ZNF536 , yang terkait dengan sistem saraf.
Keadaan iklim AP menawarkan peluang luar biasa untuk mengungkap determinan genetik yang mendorong adaptasi terhadap kondisi lingkungan yang menantang. AP menunjukkan beragam pola iklim musiman dan regional. Secara khusus, wilayah tertentu mengalami suhu musim panas yang terik yang melebihi 50°C (122°F), yang mengakibatkan lingkungan yang sangat panas dan kering (Abou-Shaara et al., 2013 ). Sebaliknya, suhu dapat anjlok hingga beku selama musim dingin, terutama di wilayah utara. Hebatnya, meskipun menghadapi tantangan lingkungan yang berat, ayam lokal tidak hanya bertahan hidup tetapi juga hidup berdampingan secara harmonis dengan manusia dalam kondisi yang keras ini (Fathi, Al-Homidan, Motawei, et al., 2017 ; Soliman et al., 2016 ). Tanpa sumber statistik yang komprehensif tentang ukuran populasi mereka atau jumlah peternakan yang membesarkan mereka, nasib sumber daya genetik ayam lokal di AP masih belum pasti. Fathi, Al-Homidan, Motawei, et al. ( 2017 ) menyoroti bahwa ras asli berada pada risiko kepunahan akibat erosi genetik yang disebabkan oleh perkawinan silang yang tidak terkendali dengan ras eksotik, yang semakin diperburuk oleh praktik manajemen yang tidak memadai.
Di sini, kami menyelidiki ayam lokal dari AP yang bertujuan untuk menjembatani kesenjangan pengetahuan dalam keragaman genom ayam dan adaptasi di wilayah tersebut. Meskipun ada bukti observasional bahwa populasi ayam lokal beradaptasi dengan baik terhadap lingkungan yang menantang, mekanisme genetik yang mendorong adaptasi ini masih kurang dipelajari (Muchadeyi & Dzomba, 2017 ). Selain itu, upaya minimal telah dilakukan untuk membedah keragaman genetik dan susunan ayam lokal di wilayah AP.
BAHAN DAN METODE
Pengambilan sampel dan pengurutan
Urutan genom dari 156 sampel dari 15 populasi ayam asli dari berbagai wilayah geografis yang mencakup zona iklim yang berbeda (Tabel 1 dan Tabel S1 ) disertakan dalam penelitian ini. Sepuluh dari populasi ini telah dipelajari sebelumnya, sementara lima populasi adalah data baru. Ini termasuk Fayoumi ( n = 10), yang berasal dari Mesir, Black feather (BL-KFU; n = 13), dan Brown feather (BR-KFU; n = 15), yang dibesarkan di King Faisal University Research and Training Station di wilayah timur Kerajaan Arab Saudi. Burung Fayoumi mencakup delapan betina dan dua jantan, yang dipilih secara acak dari kawanan yang berjumlah 700 ekor burung. Burung BL-KFU dan burung BR-KFU masing-masing mencakup 10 betina dan tiga jantan, yang dipilih secara acak dari kawanan yang berjumlah 500 ekor burung dan 1000 ekor burung. Selain itu, kami mengambil sampel burung asli di tingkat peternakan untuk dua populasi asli. Burung-burung ini adalah Buqyiq (BU-VI; n = 13) dari kota Buqyiq di Arab Saudi dan burung Oman ( n = 20) dari Negara Bagian Barka di Oman. BU-VI mencakup 10 betina dan tiga jantan. Mereka diambil sampelnya dari satu peternakan dengan populasi ayam lokal sebanyak 100 ekor. Semua ayam Oman adalah betina. Data genom dari sampel yang tersisa diunduh dari basis data genom publik (Tabel S1 ). Koordinat sistem penentuan posisi geografis dicatat untuk populasi baru (lihat Tabel S1 ). Untuk genom yang baru diurutkan, sampel darah dikumpulkan dari vena sayap dalam tabung EDTA. DNA diekstraksi menggunakan Qiagene DNeasy Blood and Tissue Kits mengikuti protokol pabrik pembuatnya ( https://www.qiagen.com/ca/resources/download.aspx?id=63e22fd7-6eed-4bcb-8097-7ec77bcd4de6&lang=en ) dan disuspensikan kembali pada konsentrasi akhir 50 ng/μl. DNA diurutkan di NEOGEN ( https://www.neogen.com ) di Inggris untuk pengurutan genom lengkap pada cakupan 20x pada platform HiSeq Illumina.
| Populasi | Ukuran sampel | Kisaran suhu di lokasi pengambilan sampel tahun 2020–2021 https://www.worldclim.org/ | Ketinggian (m) | Rata-rata curah hujan (mm) | Klasifikasi ekologi ( https://weatherandclimate.com/countries ) | Wilayah geografis (negara) | Referensi | Nomor akses proyek ENA | |
|---|---|---|---|---|---|---|---|---|---|
| Minimum (°C) | Maksimum (°C) | ||||||||
| Populasi iklim panas | |||||||||
| Bahasa Oman | 20 | 19 | 42 | tahun 1200 | 80–100 | Gurun subtropis/kering | Oman—Negara Barka | Urutan baru | PRJEB77573 |
| orang Arab Saudi | 5 | 17 | 43 | 154 | 74 | Gurun subtropis/kering | Arab Saudi | Lawal dkk. ( 2018 ) | PRJNA453469 |
| Fayoumi | 10 | Sampel populasi Mesir di Alhufuf (Arab Saudi) | Urutan baru | PRJEB77573 | |||||
| BR-KFU | 15 | Alhufuf (Arab Saudi) | |||||||
| BL-KFU | 13 | ||||||||
| BU-VI | 12 | 90 | |||||||
| Populasi iklim dingin | |||||||||
| Pelantun | 9 | -25 | 20 | tahun 1652 | >1000 | Dingin/sedikit lembab | Quebec (Kanada) | Xu dan kawan-kawan ( 2021 ) | PRJNA720223 |
| Dulong | 10 | -6 | 16 | 3000 | >3000 | Daerah tropis yang dingin di dataran tinggi | Sabuk Ngarai Alpen di Pegunungan Hengduan Utara–Selatan (Republik Rakyat Tiongkok) | Wang dkk. ( 2020 ) | PRJNA559932 |
| Tibet | 5 | -7 | 23 | 3650 | – | Dingin/kering | Dataran Tinggi Tibet (Republik Rakyat Tiongkok) | Zhang dkk. ( 2016 ) | PRJNA309581 |
| Populasi Ethiopia di daerah dingin | |||||||||
| Alfa Midir | 9 | 1 | 19 | 3451 | >1000 | Tropis dingin/sub-lembab | Pegunungan Amhara Tengah (Ethiopia) | Gheyas dan kawan-kawan ( 2022 ) | PRJEB39275 |
| Gafera | 10 | 5 | 25 | tahun 2592 | 900–1000 | ||||
| Negasi Amba | 10 | 3 | 20 | 3071 | 900–1000 | ||||
| Populasi Ethiopia di daerah hangat | |||||||||
| Gesses | 10 | 11 | 33 | tahun 1546 | >900 | Daerah tropis hangat/semi-kering | Lembah Amhara Barat (Etiopia) | Gheyas dan kawan-kawan ( 2022 ) | PRJEB39275 |
| Pelukan | 9 | 15 | 37 | 979 | >900 | Afar Timur (Ethiopia) | |||
| Anak kecil | 9 | 11 | 33 | tahun 1418 | >900 | Daerah tropis hangat/kering | Lembah Amhara Barat (Etiopia) | ||
Pemetaan dan pemanggilan varian
Pembacaan sekuens genom secara keseluruhan dipetakan ke rakitan genom referensi “GRCg6a”, menggunakan algoritma bwa-mem dari burrows-wheeler aligner versi 0.7.17 (Li & Durbin, 2009 ). Pembacaan yang dipetakan diurutkan berdasarkan koordinat dan duplikat PCR ditandai dan dihapus menggunakan opsi SortSam dan MarkDuplicates dari picard versi 2.18.29 ( https://broadinstitute.github.io/picard/ ), masing-masing. Setiap sampel kemudian dikenakan kalibrasi ulang skor kualitas dasar menggunakan “situs yang diketahui” dbSNP ( http://ftp.ensembl.org/pub/release-104/variation/vcf/gallus_gallus/ ). Proses ini ditujukan untuk menghilangkan kesalahan atau ketidakselarasan selama pengurutan dan mengidentifikasi varian yang diketahui. Polimorfisme nukleotida tunggal (SNP) dipanggil menggunakan algoritma HaplotypeCaller dari genome analysis toolkit (gatk ) versi 4.2.2.0 pada mode GVCF (McKenna et al., 2010 ) pada setiap sampel. Tempat kerja untuk setiap kromosom diterapkan menggunakan algoritma GenomicsDBImport . Genotipe gabungan dilakukan untuk mengidentifikasi varian pada semua sampel secara bersamaan diikuti oleh penyaringan keras menggunakan algoritma VariantFiltration untuk pemanggilan SNP berkualitas tinggi. Selanjutnya, varian autosomal bi-allelic yang lolos langkah penyaringan varian dipilih untuk analisis hilir.
Keanekaragaman genom dan struktur populasi
Beberapa metode digunakan untuk mengeksplorasi keragaman genetik dan struktur populasi. vcftools versi 0.1.14 (Danecek et al., 2011 ) digunakan untuk estimasi keragaman genetik pada tingkat genom, termasuk diferensiasi populasi berpasangan ( F ST ) dan heterozigositas. Rata-rata genom F ST dihitung menggunakan jendela 20-kb dengan langkah geser 10 kb. Analisis kontrol kualitas dilakukan pada SNP autosomal yang disebut untuk tujuan analisis keragaman genetik dan struktur populasi menggunakan plink 1.9 (Chang et al., 2015 ). SNP dengan frekuensi alel minor ≤ 0,05 (4.289.890 SNP), dan tingkat panggilan genotipe ≤ 95% dikecualikan (4.143 SNP). Untuk analisis keragaman genetik, SNP berpasangan dengan ketidakseimbangan hubungan tinggi (koefisien korelasi kuadrat r 2 > 0,1) selanjutnya dikecualikan (17.237.390 SNP). Analisis komponen utama dilakukan dengan 3.374.709 SNP yang tersisa menggunakan opsi –pca dalam plink 1.9 dan diplot menggunakan paket r tidyverse (Wickham et al., 2019 ). Program admixture versi 1.3.0 (Alexander et al., 2009 ), untuk hingga 12 klaster leluhur yang disimpulkan ( K ), digunakan untuk menilai proporsi leluhur genom yang optimal dalam populasi ayam yang dianalisis.
Tanda-tanda analisis seleksi
Kami melakukan empat tes di semua populasi untuk mendeteksi tanda-tanda kandidat seleksi positif termasuk heterozigositas gabungan Hp (Rubin et al., 2010 ), skor haplotipe terintegrasi iHS (Voight et al., 2006 ), diferensiasi populasi F ST (Akey et al., 2002 ), dan homozigositas haplotipe lintas populasi yang diperluas XP-EHH (Sabeti et al., 2007 ). Kami menggunakan jendela geser 20-kb dengan langkah 10-kb di semua metode dengan minimal 20 SNP per jendela. Nilai Hp dan F ST distandarisasi ( ZHp dan ZF ST ) untuk memastikan konsistensi di semua analisis. Analisis iHS dan XP-EHH dilakukan menggunakan perangkat lunak HAPBIN (Ahbara et al., 2022 ; Maclean et al., 2015 ) setelah menghilangkan SNP dengan genotipe yang hilang. Analisis iHS dan XP-EHH dilakukan untuk SNP individual, dan kemudian nilai rata-rata dihitung dalam jendela untuk metrik iHS standar ( iHS _std) dan XP-EHH ( XP-EHH _std).
Identifikasi gen dan anotasi fungsional
Kami menggunakan rakitan genom referensi “GRCg6a”, rilis 110 basis data dbSNP, (2023), dan “VEP” Ensembl (McLaren et al., 2016 ) untuk posisi SNP dan identifikasi gen. Ambang batas untuk deteksi outlier melibatkan pemilihan persentil terendah ekstrem 0,001 dari ZHp , dan persentil tertinggi ekstrem 0,001 untuk nilai iHS , ZF ST , dan XP-EHH , dari distribusi empiris masing-masing.
Wilayah yang diidentifikasi kemudian dikonsolidasikan menggunakan bedtools v.2.25.0. Gen yang tumpang tindih dengan wilayah kandidat diidentifikasi berdasarkan basis data 108 Ensembl Genes menggunakan alat Ensembl BioMart daring ( http://www.ensembl.org/biomart ). Dua daftar terpisah dibuat: (i) gen yang tumpang tindih atau dalam tanda tangan wilayah seleksi menggunakan analisis ZHp dan iHS dalam populasi ; dan (ii) gen yang tumpang tindih atau dalam tanda tangan seleksi yang diidentifikasi oleh perbandingan berpasangan F ST dan XP-EHH antar populasi . Kami menggunakan perangkat lunak Database for Annotation, Visualization, and Integrated Discovery (DAVID) versi 6.8 ( https://david.ncifcrf.gov/summary.jsp ) untuk meneliti daftar gen untuk pengayaan gen yang signifikan yang terkait dengan kategori fungsional tertentu. Analisis DAVID mencakup berbagai kategori anotasi, seperti Gene Ontology (GO), Proses Biologis, dan Fungsi Molekuler GO, untuk mengidentifikasi proses dan fungsi biologis yang diperkaya dalam daftar gen. Ambang signifikansi nilai p yang dikoreksi Benjamini sebesar 0,05 digunakan untuk menentukan pengayaan statistik. Lebih jauh, kami menyelidiki peran biologis setiap gen yang dianotasi melalui penelusuran literatur ekstensif yang menggabungkan informasi dari berbagai spesies.
HASIL
Pengurutan dan analisis varian
Urutan genom sampel ayam dari Arab Saudi, Oman, dan Mesir menghasilkan rata-rata 221–480 juta pembacaan urutan berpasangan. Cakupan genom rata-rata untuk urutan tersebut berkisar antara 16,98× hingga 57,46× (lihat Tabel S2 ). Pembacaan tersebut diselaraskan dengan genom referensi ayam (GRCg6a), dan analisis gabungan dari semua sampel kemudian dilakukan. Analisis ini mengungkap sekitar 25 juta SNP berkualitas tinggi untuk 15 populasi. Menggabungkan semua populasi menemukan bahwa 38,25% dari seluruh set SNP (9,5 juta dari 24,9 juta) adalah baru menurut dbSNP rilis 110 (2023). Di antara populasi yang diperiksa, Chantecler memiliki persentase terendah (12,4%) SNP baru, dan populasi AP BL-KFU, BR-KFU, dan BU.VI memiliki yang tertinggi masing-masing 22,31%, 24,44%, dan 24,73% (Tabel S3 ). Kepadatan SNP yang diamati di seluruh kromosom diilustrasikan dalam Tabel S4 . Kepadatan rata-rata SNP di seluruh autosom adalah 26,49 SNP/kb. Kromosom 22 menunjukkan kepadatan SNP terendah, sedangkan kromosom 31 menunjukkan kepadatan tertinggi, diikuti oleh kromosom 33.
Struktur dan diferensiasi populasi
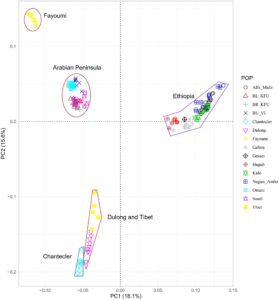
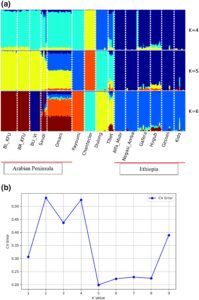
Struktur populasi diperiksa menggunakan analisis komponen utama, campuran dan analisis diferensiasi genetik ( FST ) . Komponen utama (PC)1, yang mencakup 18,1% variasi, secara jelas membedakan semua populasi Ethiopia dari populasi yang tersisa, sementara PC2, yang menjelaskan 15,6% dari total variasi, memisahkan populasi AP dan Fayoumi dari Chantecler, serta sebagian besar populasi Ethiopia dari Dulong dan Chantecler (Gambar 1 ). Populasi Oman dan Arab Saudi terletak pada posisi perantara pada Gambar 1. Analisis campuran ( K = 5) mendukung satu kumpulan gen leluhur untuk populasi Arab Saudi, sedangkan populasi Oman tampak jauh lebih bercampur (Gambar 2 ). Populasi lain menunjukkan susunan genetik yang berbeda, seperti yang ditunjukkan oleh latar belakang genetik mereka yang berbeda. Peta panas analisis F ST berpasangan (Tabel 2 ) menggambarkan bahwa diferensiasi populasi terendah diamati antara populasi Ethiopia Alfa Midir dan Negasi Amba ( F ST = 0,009), Gafera dan Gesses ( F ST = 0,022), Gesses dan Kido ( F ST = 0,022), dan Gafera dan Kido ( F ST = 0,024), serta antara populasi Cina Dulong dan Tibet ( F ST = 0,035). Sebaliknya, diferensiasi populasi tertinggi ditemukan antara Chantecler dan Fayoumi ( F ST = 0,349). Secara umum, ada diferensiasi genetik sedang antara populasi kecuali untuk Fayoumi dan Chantecler, yang menunjukkan nilai F ST tertinggi dengan populasi lainnya.
TABEL 2. Fiksasi populasi berpasangan ( FST ) di antara populasi ayam.

Mendeteksi sinyal sapuan selektif di seluruh genom
Empat pendekatan berbeda digunakan dalam uji seleksi untuk mengidentifikasi adaptasi termotoleransi (stres panas dan dingin). Pertama-tama kami bertujuan untuk mengidentifikasi kandidat daerah genom yang berpotensi di bawah seleksi positif untuk termotoleransi dalam setiap populasi secara terpisah. Daerah genom spesifik populasi dibandingkan di seluruh kelompok untuk mengevaluasi tumpang tindih antara populasi iklim panas (AP dan Fayoumi) dan populasi iklim dingin (Chantecler, Dulong, dan Tibet). Tujuannya adalah untuk mengidentifikasi kandidat daerah yang menunjukkan diferensiasi signifikan dalam frekuensi alel (menggunakan metode ZF ST ) atau dalam pola ketidakseimbangan hubungan (menggunakan metode XP-EHH ). Jendela kandidat diidentifikasi melalui analisis tanda tangan seleksi dengan persentil ekstrem 0,001, di mana ambang batas ditetapkan pada F ST standar ( ZF ST ) >3,24–6,24, dan XP-EHH standar absolut (| XP-EHH _std|) >3,24–4,11.
Wilayah kandidat dan gen yang terkait dengan adaptasi terhadap kondisi ekstrem dalam populasi
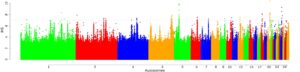
Kami menganalisis 81.007–95.110 dan 95.101–95.122 jendela di seluruh populasi dalam analisis ZHp dan iHS , masing-masing (Tabel S5 ). Sapuan ZHp yang signifikan terdeteksi di seluruh autosom genom ayam, kecuali pada kromosom 16, 19, 20, 21, 22, 25, 28, dan 31 (Gambar 3 dan Tabel S6 ). iHS diamati di seluruh autosom kecuali kromosom 25, 27, dan 31 (Gambar 4 , Tabel S7 ). Di seluruh populasi AP di bawah tekanan panas (Oman, Arab Saudi, dan Fayoumi), jendela ini secara kolektif mencakup 169 gen ( ZHp ) dan 172 gen ( iHS ) Tabel 3 . Hasil terperinci disediakan dalam Tabel S8a–n untuk ZHp dan S9a–n untuk iHS , serta dalam Gambar S1a–l dan S2a–n . Gen kandidat menunjukkan fungsi biologis yang berbeda, khususnya terkait dengan toleransi panas, respons stres, imunitas, dan sistem saraf (Tabel S10 ). Analisis istilah GO dan jalur Kyoto Encyclopedia of Genes and Genomes (KEGG) yang terkait dengan gen-gen ini memberikan wawasan tambahan ke dalam potensi pentingnya mereka untuk jalur biologis yang berbeda, termasuk toleransi panas, pensinyalan adrenergik dalam kardiomiosit, respons terhadap bakteri, dan fosforilasi protein (Tabel S11 dan S12 ). Misalnya, RYR2 terlibat dalam takikardia ventrikel polimorfik yang diinduksi stres, dan yang menarik, itu terdeteksi pada populasi Tiongkok dalam kondisi dingin (Wang et al., 2015 ). LDB2 berkontribusi pada pembentukan pembuluh darah (Javerzat et al., 2009 ). Selain itu, APP , NTN3 , dan PUF60 dikaitkan dengan respons apoptotik terhadap stres (Gu et al., 2020 ), sementara CNTNAP2 berperan dalam proses sistem saraf (von Holdt et al., 2023 ) terdeteksi dalam populasi BL-KFU. Dalam populasi AP, ALX4 dan PLA2G15 , yang berperan dalam deposisi lemak dan proses metabolisme asam lemak (McManus et al., 2022 ; Zhang et al., 2016 ), terdeteksi bersama dengan COL6A2 , gen kunci dalam sintesis kolagen (Fleming et al., 2017 ; Wang et al., 2016 ). Menariknya, COL6A2Gen tersebut juga terdeteksi di wilayah genom yang menunjukkan sinyal ZHp negatif yang substansial (−4,67) di Gesses dan populasi Ethiopia lainnya, yang tinggal di lokasi tropis hangat/semi-kering. Lebih jauh, ORAOV1 dikaitkan dengan resistensi terhadap stres oksidatif, FGF19 membantu penyesuaian metabolisme selama puasa, dan PLCE1 berpotensi memengaruhi kandungan lemak intramuskular yang lebih tinggi.

| Bahasa Inggris CHR | Mulai (bp) | Akhir (bp) | Metode | Skor tertinggi | Gen dan Fungsinya |
|---|---|---|---|---|---|
| 5 | Nomor telepon 17 070 001 | 17.140.000 | Bahasa Indonesia: IHS | 8.24 | ORAOV1 terlibat dalam resistensi terhadap stres oksidatif (Togashi et al., 2014 ) |
| FGF19 memfasilitasi penyesuaian metabolisme selama puasa (Degirolamo et al., 2016 ) | |||||
| 6 | 20 920 001 | 20.940.000 | Bahasa Indonesia: IHS | 8.42 | PLCE1 terlibat dalam metabolisme (Li et al., 2022 ) |
| 1 | Nomor telepon 102 900 001 | Nomor telepon 102 920 001 | ZHp | -4,28 | APP mengatur apoptosis sel (Gu et al., 2020 ) |
| 2 | Nomor telepon 149 040 001 | Nomor telepon 149 060 001 | ZHp | -3,90 | PUF60 dikaitkan dengan apoptosis (Dong et al., 2019 ) |
| 5 | 21 340 001 | 21.390.000 | ZHp | -3,98 | ALX4 dikaitkan dengan penumpukan lemak (McManus et al., 2022 ) |
| 11 | 50 001 | 70.000 orang | ZHp | -3,76 | PLA2G15 dikaitkan dengan proses metabolisme asam lemak (Zhang et al., 2016 ) |
| 3 | Nomor telepon 37 070 001 | Nomor telepon 37 090 001 | ZHp | -3,94 | RYR2 memiliki peran dalam takikardia ventrikel polimorfik yang disebabkan oleh stres (Wang et al., 2015 ) |
| 2 | 53 320 001 | 53.340.000 | ZHp | -3,96 | Peran CNTNAP2 adalah dalam proses sistem saraf dan mungkin memainkan peran penting dalam bagaimana individu menyesuaikan diri dengan lingkungan baru (von Holdt et al., 2023 ) |
Tiga belas dari 133 gen yang terdeteksi di ZHp dan iHS mungkin terkait dengan adaptasi terhadap iklim dingin. Gen yang relevan dengan neurobehavioral, CNTNAP2 , diidentifikasi pada populasi Chantecler dengan sinyal tertinggi ( ZHp = −3.76) dalam wilayah 340 kb. SRBD1 (protein pengikat RNA S1 1) diidentifikasi di seluruh populasi yang tinggal di lingkungan dingin. Ini telah dikaitkan kuat dengan induksi apoptosis sel (Ung et al., 2017 ). Pada populasi Tiongkok, skor ZHp terendah diamati pada Kromosom 24, yang berisi 10 gen berbeda ( BCO2 , TEX12 , IL18 , SDHD , PIH1D2 , DLAT , C11ORF52 , CRYAB , HSPB2 , dan DIXDC1 ). Gen STK38L diidentifikasi pada populasi Tiongkok ( analisis iHS ) dan terlibat dalam pembelahan dan morfologi sel saraf (Goldstein et al., 2010 ).
Kandidat potensial dan gen untuk adaptasi stres panas membandingkan populasi dari kondisi panas-kering dan dingin
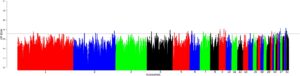
Kami menganalisis 94.610 hingga 95.035 dan 94.025 hingga 95.185 jendela di seluruh populasi dengan analisis ZF ST dan XP-EHH , masing-masing (Tabel S13 ). Untuk menyederhanakan analisis, kami hanya mempertimbangkan jendela kandidat yang tumpang tindih pada persentil ekstrem 0,001 dari distribusi empiris. Ini termasuk 392 dan 336 daerah sapuan untuk ZF ST dan XP-EHH , masing-masing, di seluruh perbandingan. Khususnya, sapuan ZF ST yang signifikan terdeteksi di seluruh autosom, kecuali pada kromosom 16, 17, 18, 22, 23, 24, 25, 26, 27, 28, 31, 32, dan 33 (Gambar 5 ). Sapuan XP-EHH diamati di seluruh autosom kecuali kromosom 3 (Gambar 6 ). Jendela-jendela ini secara kolektif mencakup 226 gen ( analisis ZF ST ) dan 275 gen ( analisis XP-EHH ) untuk sembilan perbandingan berpasangan populasi dari kondisi panas-kering (Oman, Arab Saudi, dan Fayoumi) dibandingkan dengan lingkungan dingin (Chantecler, Dulong dan Tibet) Tabel 4. Hasil terperinci disediakan dalam Tabel S14a–j dan (Gambar S3a–d dan S4a–f ) untuk ZF ST dan Tabel S15a–j dan Gambar S5a–c dan S6a–d untuk XP-EHH . Gen-gen, yang diidentifikasi menggunakan kombinasi setidaknya dua metode (Tabel 5 dan Gambar 7 ), ditemukan terkait dengan sifat adaptasi lingkungan yang berbeda, termasuk apoptosis, pengaturan tekanan darah, pengaturan pigmentasi kulit, hipoksia, pengaturan keseimbangan antara penyimpanan energi dan pengeluaran energi, metabolisme, dan respons imun. Lima istilah jalur KEGG yang terkait dengan jalur biologis yang berbeda: pematangan oosit yang dimediasi progesteron, pensinyalan adrenergik dalam kardiomiosit, jalur pensinyalan GnRH, kontraksi otot polos vaskular, dan meiosis oosit, didefinisikan untuk gen-gen ini (Tabel S16 ).

| Bahasa Inggris CHR | Mulai (bp) | Akhir (bp) | Metode | Skor tertinggi | Gen dan fungsinya | Perbandingan |
|---|---|---|---|---|---|---|
| 1 | 65 790 001 | 65.810.000 | ZF- ST-Serigala | 4.5 | Bahasa Indonesia: SOX5 | BU-VI vs. Chantecler |
| 1 | 65 900 001 | 65.920.000 | ZF- ST-Serigala | 5.44 | Bahasa Indonesia: SOX5 | Bl-KFU melawan Chantecler |
| 2 | 72 660 001 | 72.680.000 orang | ZF- ST-Serigala | 7.73 | CTNND2 | Populasi Oman vs. Tiongkok |
| 3 | 50 260 001 | 50 280 000 | ZF- ST-Serigala | 5.28 | TFB1M | Oman vs. Chantecler |
| 5 | 45 420 001 | 45.440.000 | ZF- ST-Serigala | 4.11 | KLMN | Fayoumi melawan Chantecler |
| 5 | 4 120 001 | 4.140.000 | ZF- ST-Serigala | 6.39 | METTL15P1 | Populasi Fayoumi vs. Tiongkok |
| 7 | 9 190 001 | 9.210.000 orang | ZF- ST-Serigala | 7.35 | SLC39A10 | Populasi Br-KFU vs. Tiongkok |
| 8 | 1 010 001 | 1.030.000 orang | ZF- ST-Serigala | 7.98 | VAV3 | Bl-KFU vs. populasi Tiongkok (Dulong dan Tibet) |
| 20 | Nomor telepon 6 070 001 | 6.090.000 orang | ZF- ST-Serigala | 5.35 | EYA2 | Br-KFU melawan Chantecler |
| 18 | 11 300 001 | 11.330.000 | XP-EHH | 5.34 | SUMO2 dan NUP85 | BU-VI vs. Chantecler |
| 24 | 6 290 001 | 6.310.000 orang | XP-EHH | 4.84 | C11ORF52 , CRYAB , HSPB2 , C11orf1 , FDXACB1 , dan ALG9 | Bl-KFU melawan Chantecler |
| 16 | 620 001 | 650.000 | XP-EHH | 4.58 | 5_8S_rRNA | Populasi Oman vs. Tiongkok (Dulong dan Tibet) |
| 24 | 6 290 001 | 6.320.000 orang | XP-EHH | 4.55 | C11ORF52 , CRYAB , HSPB2 , C11orf1 , FDXACB1 , dan ALG9 | Oman vs. Chantecler |
| 14 | 250 001 | 280.000 | XP-EHH | 3.90 | CPPED1 | Fayoumi melawan Chantecler |
| 12 | 14 590 001 | 14.620.000 | XP-EHH | 3.98 | SLC25A26 dan LRIG1 | Br-KFU vs. populasi Tiongkok (Dulong dan Tibet) |
| 9 | Nomor telepon 18 070 001 | 18.090.000 | XP-EHH | 4.10 | NAALADL2 | Bl-KFU vs. populasi Tiongkok (Dulong dan Tibet) |
| 28 | 5 050 001 | 5 100 000 | XP-EHH | 5.14 | PTPRS | Br-KFU melawan Chantecler |
| Krom | Daerah kromosom | Analisa | Gen | Fungsi | |
|---|---|---|---|---|---|
| 4 | 75.820.001–75.850.000 | BL-KFU vs. Chantecler ( ZF ST ), Br-KFU vs. Chantecler ( ZF ST ), Oman vs. Chantecler ( ZF ST ), populasi Oman vs. Tiongkok ( ZF ST ) | Populasi Cina ( ZHp ), Br-KFU ( ZHp ), BU-VI ( ZHp ), dan Fayoumi ( ZHp ) | LDB2 | Terlibat dalam perkembangan otak, pembentukan pembuluh darah (Javerzat et al., 2009 ) |
| 5 | 3.490.001–3.510.000 | Fayoumi vs. Chantecler ( ZF ST ) dan Fayoumi vs. populasi Tiongkok ( ZF ST ) | BU-VI ( iHS ) | ANO3 | Terkait dengan metabolisme (Jean et al., 2015 ) |
| 6 | 17.400.001–17.430.000 | Br-KFU_vs. populasi Tiongkok ( ZF ST ), Br-KFU. vs. populasi Tiongkok ( XP-EHH ), dan Br-KFU vs. Chantecler ( ZF ST ) | Bahasa Indonesia: ITGA6 | Terkait langsung diatur oleh faktor-faktor yang dapat menginduksi hipoksia (Brooks et al., 2016 ) | |
| 7 | 6.670.001–6.710.000 | Br-KFU vs. populasi Cina ( ZF ST ), Br-KFU vs. Chantecler ( ZF ST ) | BL-KFU ( ZHp ) dan Fayoumi ( ZHp ) | COL6A2 | Terlibat dalam biosintesis kolagen dan modifikasi enzim (Fleming et al., 2017 ; Wang et al., 2016 ) |
| 14 | 280.001–310.000 | Br-KFU vs. Chantecler ( XP-EHH ), Fayoumi vs. Chantecler ( XP-EHH ), dan Oman vs. Chantecler ( XP-EHH) ) | BU-VI ( iHS ), dan Fayoumi ( iHS ) | CPPED1 | Terkait dengan proses sistem kekebalan tubuh (Bentz et al., 2019 ; Chen et al., 2022 ) |
| 15 | 3.390.001–3.420.000 | BL-KFU melawan Chantecler ( ZF ST ) | Populasi Tiongkok ( ZHp ) | FZD10 | Terlibat dalam pengaturan pigmentasi kulit, aspek penting perlindungan terhadap radiasi matahari. (Nie et al., 2022 ) |
| 18 | 3 830 001–3 870 000 | BL-KFU vs. populasi Cina ( XP-EHH ), dan BL-KFU vs. Chantecler ( ZF ST ) | CASZ1 | Memainkan peran penting dalam mengatur tekanan darah (Simino et al., 2014 ) | |
| 19 | 9 350 001–9 410 000 | BL-KFU vs. Chantecler ( ZF ST ), dan BU-VI vs. Chantecler ( XP-EHH ) | MSI2 | Terlibat dalam proses apoptosis (Bennett et al., 2016 ) | |
| 23 | 5.950.001–5.980.000 | Oman vs. Chantecler ( XP-EHH) ) | BR-KFU ( iHS ). | PNRC2 | Terlibat dalam mengatur keseimbangan antara penyimpanan energi dan pengeluaran energi (Sorbolini et al., 2017 ) |
| SRSF10 | Mungkin terlibat dalam meningkatkan kelangsungan hidup sel selama kondisi stres (Zhou et al., 2014 ) | ||||
| 24 | 6 290 001–6 320 000 | BU_VI vs. Chantecler ( XP-EHH ), Bl-KFU vs. Chantecler ( XP-EHH ), Br-KFU vs. Chantecler ( XP-EHH ), dan Oman vs. Chantecler ( XP-EHH) ) | Penyanyi ( iHS ) | Bahasa Inggris ALG9 | Terkait dengan metabolisme lipid (Iqbal et al., 2023 |
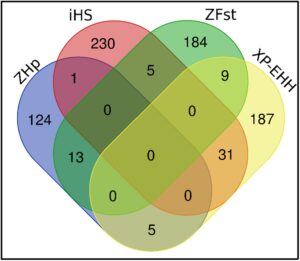
Puncak paling signifikan ada pada kromosom 2, 4, 7, 8, 11, dan 19 dalam analisis ZF ST (semua sembilan perbandingan). Daerah kandidat ini meliputi VAV3 yang terkait dengan hipoksia, yang sebelumnya dilaporkan pada ayam Ethiopia (Scheinfeldt et al., 2012 ). Gen menarik lainnya adalah: NTNG1 , yang terlibat dalam sekresi, secara signifikan diatur naik oleh stres panas pada garis sel hepatoseluler ayam White Leghorn jantan (Sun et al., 2015 ); VSTM2A terkait dengan perilaku (Wang et al., 2023 ); SMPD3 dan EYA2 terkait dengan kinerja pertumbuhan (Bounas et al., 2023 ; Feng et al., 2023 ); DYNLL2 untuk respons imun (Ogada et al., 2023 ); SLC39A10 untuk metabolisme (Zhu et al., 2019 ); COL18A1 untuk biosintesis kolagen dan modifikasi enzim (Wang et al., 2016 ); dan tiroglobulin TG yang diketahui memengaruhi kemampuan untuk mengumpulkan lemak intramuskular (Kostusiak et al., 2023 ). Salah satu gen yang terkait dengan sirkulasi darah adalah SOX5 , yang sangat penting dalam mengarahkan aliran darah ke kulit dan memfasilitasi pertukaran panas selama suhu tinggi (Hester et al., 2015 ).
Sedangkan untuk hasil XP-EHH , kami temukan dalam region genom kandidat SLC25A26 , LRIG1 , dan JADE3 . Juga hadir heat shock protein family B (small) member 2 ( HSPB2 ) dan crystallin α B ( CRYAB ) yang terlibat dalam toleransi stres, integritas sel otot, dan kelangsungan hidup sel terhadap stres panas (Du et al., 2022 ; Nakagawa et al., 2001 ; Wang et al., 2016 ), ALG9 (metabolisme lipid) (Iqbal et al., 2023 ), PTPRS (jalur pensinyalan insulin dan inflamasi) (Samblas et al., 2018 ), dan NUP85 (respons stres) (Zhu et al., 2017 ).
DISKUSI
Keanekaragaman dan struktur populasi
Studi ini merupakan analisis urutan genom utuh skala besar pertama dari ayam lokal dari AP, yang meliputi bagian timur Arab Saudi dan Oman. Tujuan utamanya adalah untuk menilai keragaman genetik dan lanskap adaptif genom ayam lokal dari AP, sebagai respons terhadap tantangan termal ekstrem (iklim panas dan kering), serta membandingkan populasi ini dengan populasi ayam lain yang dikenal karena adaptasinya terhadap kondisi lingkungan dingin yang keras.
Kami menggunakan referensi genom ayam GRCg6a untuk memanggil SNP di seluruh genom untuk 156 ayam, mengidentifikasi 24,9 juta SNP, yang 38,24% di antaranya baru. Penelitian sebelumnya oleh Lawal et al. ( 2018 ) dan Gheyas et al. ( 2022 ) melaporkan 10 juta (16% baru) dan 19,5 juta (29% baru) SNP pada ayam domestik Ethiopia, masing-masing. Penjelasan yang mungkin untuk perbedaan ini mencakup versi genom referensi ayam yang berbeda, jumlah sampel, dan perbedaan dalam metrik filter dan cakupan genom. Kepadatan SNP rata-rata, dengan nilai 26,49 SNP/kb, melampaui kepadatan yang dilaporkan sebelumnya sebesar 15 SNP/kb pada populasi ayam eksperimental dan komersial berdasarkan genom referensi galGal5 (Gheyas et al., 2015 ). Kepadatan SNP pada populasi ayam asli Ethiopia, menggunakan galGal6 sebagai genom referensi, adalah sekitar 19 SNP/kb dalam studi Gheyas et al. ( 2022 ). Perbedaan tersebut menunjukkan bahwa peningkatan kualitas perakitan genom mungkin berkontribusi terhadap peningkatan kepadatan SNP yang diamati dalam studi kami.
Analisis PC dan campuran menunjukkan diferensiasi genetik antara populasi dan memisahkan sebagian besar dari mereka berdasarkan daerah asal geografis mereka. Kedekatan yang relatif diamati antara AP dan ayam Fayoumi yang mendukung warisan genetik bersama antara kedua garis ini, yang sangat beradaptasi dengan stres panas, tidak seperti populasi lainnya. Menariknya, ayam Chantecler ditemukan relatif dekat dengan populasi Cina lokal (Dulong dan Tibet) meskipun asal geografis mereka sangat berbeda, yang dipisahkan menjadi kelompok yang berbeda berdasarkan analisis campuran. Hasil ini menunjukkan bahwa sebagian besar populasi yang kami pelajari mungkin tidak diperdagangkan atau secara historis bercampur dengan yang lain untuk waktu yang lama. Pengecualiannya mungkin adalah populasi Oman, yang menunjukkan tingkat campuran yang tinggi dari latar belakang genetik yang berbeda, mungkin warisan dari beberapa introduksi di sepanjang pantai timur AP.
Gen yang diseleksi secara positif berhubungan dengan toleransi panas
Iklim gurun yang keras di AP, yang dicirikan oleh suhu yang sangat panas dan kondisi kering akibat radiasi matahari yang intens, menghadirkan tantangan yang signifikan bagi organisme hidup, termasuk ayam. Sebagai respons terhadap kondisi ekstrem ini, ayam telah mengembangkan berbagai strategi untuk mempertahankan suhu tubuh mereka. Strategi ini melibatkan penggunaan radiasi yang ditingkatkan, pembuangan panas konvektif, dan pendinginan evaporatif melalui tekanan darah diastolik mereka. Khususnya, ayam disebut sebagai “hewan yang benar-benar terengah-engah”, yang menunjukkan bahwa mereka dapat melepaskan panas internal dengan menguapkan kelembapan melalui paru-paru dan kantung udara mereka. Mekanisme terengah-engah ini berfungsi sebagai alat penting untuk mengatur suhu tubuh dalam menghadapi panas yang tak henti-hentinya (Collier & Gebremedhin, 2015 ; Lara & Rostagno, 2013 ; Mutaf et al., 2009) ). Oleh karena itu, dengan meneliti hasil dari empat metode yang berbeda untuk mendeteksi tanda tangan positif kandidat seleksi dan menyelidiki peran gen dalam wilayah ini, kita mungkin dapat menghubungkannya dengan strategi termotoleransi tertentu.
Dalam studi ini, kami mengidentifikasi beberapa gen yang terkait dengan perkembangan sistem kardiovaskular dan pembentukan pembuluh darah ( RYR2 , LDB2 , SOX5 , dan FHOD3 ) yang mungkin telah mengalami seleksi positif pada populasi ayam dari AP. Contoh penting termasuk RYR2 , yang dikenal karena hubungannya dengan takikardia ventrikel polimorfik yang diinduksi stres dan yang mungkin terkait dengan adaptasi terhadap dataran tinggi pada ayam Tibet (Wang et al., 2015 ). Gen lain dengan fungsi penting adalah LDB2 , yang memainkan peran penting dalam pembentukan pembuluh darah (Javerzat et al., 2009 ), dan SOX5 , yang dikenal pada ayam Leghorn untuk mengalihkan aliran darah ke kulit, memfasilitasi pertukaran panas (Hester et al., 2015 ). Yang juga layak disebutkan adalah FHOD3 , yang termasuk dalam keluarga protein formin, yang memiliki peran penting dalam perkembangan jantung (Rosado et al., 2014 ). FHOD3 diekspresikan secara menonjol di jantung dan memainkan peran penting dalam miofibrilogenesis, khususnya dalam pematangan miofibril (Kan-o et al., 2012 ). Semua gen ini mungkin terkait erat dengan pengaturan aliran darah dan mekanisme pendinginan evaporatif.
Kami juga telah mengidentifikasi pengayaan jalur fungsional yang signifikan setelah analisis KEGG ( p < 0,05). Ini mencakup jalur pensinyalan adrenergik dalam kardiomiosit. Temuan ini sangat penting karena pensinyalan adrenergik adalah pengatur utama fungsi jantung, yang memungkinkan burung-burung ini untuk secara dinamis menyesuaikan detak jantung dan respons kardiovaskular mereka sebagai respons terhadap perubahan kondisi lingkungan yang mereka hadapi. Empat gen ( PRKCD , BRAF , ADCY7 , dan ADCY6 ), bagian dari jalur kontraksi otot polos vaskular, juga diidentifikasi. Jalur ini terlibat dalam pengendalian fungsi sistolik dan diastolik yang merespons berbagai tuntutan fisiologis (Schaub et al., 2006 ) . Menariknya, beberapa hasil ini sejalan dengan penelitian sebelumnya yang dilakukan pada ayam lokal Arab Saudi oleh Tian et al. ), yang juga menekankan peran sistem adrenergik dan muskarinik dalam mengendalikan denyut jantung dan fungsi jantung dalam menanggapi berbagai tantangan fisiologis.
Kehadiran gen yang terkait dengan apoptosis merupakan temuan signifikan dalam studi ini. Stres panas dapat menyebabkan cedera seluler di hati, jantung, ginjal, dan sistem saraf pusat, yang menekankan pentingnya memahami dasar genetik regulasi apoptosis sebagai respons terhadap stresor lingkungan. Apoptosis adalah proses kematian sel yang sangat diatur dan dikendalikan yang penting untuk menjaga stabilitas internal pada organisme hidup dan memfasilitasi adaptasi terhadap lingkungan sekitar (Elmore, 2007 ). Dalam konteks kondisi lingkungan AP yang menantang, temuan ini membawa signifikansi khusus. Salah satu gen yang diidentifikasi, SRBD1 , diketahui berperan dalam pertumbuhan sel dan regulasi apoptosis (Mizuki, 2010 ). Itu terkait dengan adaptasi terhadap lingkungan terkering dan terhangat dari populasi burung tit besar liar (Stonehouse et al., 2024 ). Selain itu, studi lain telah menekankan keberadaan gen yang terlibat dalam diferensiasi sel dan apoptosis pada sel saraf, seperti APP dan PUF60 pada ayam kampung Cina (Gu et al., 2020 ). Gen SLC26A8 juga dicatat, berpotensi memainkan peran dalam regulasi sel atau apoptosis (Dirami et al., 2013 ). Temuan penting lainnya adalah referensi ke gen protein terkait kematian ( DAP ), yang dikaitkan dengan regulasi miogenesis, apoptosis, dan perkembangan kerangka pada ayam (Tesseraud et al., 2021 ). Menariknya, kami juga mengidentifikasi jalur pengayaan yang terkait dengan apoptosis pada populasi Chantecler dan Chinese yang hidup di lingkungan dingin.
Kehadiran dan fungsi pigmen melanin (kulit, bulu) sangat penting untuk melindungi hewan dari efek merusak radiasi matahari. Pensinyalan Wnt sangat penting dalam mengatur stratifikasi epidermis, penting untuk menjaga kesehatan dan fungsi kulit. Dalam konteks penelitian ini, tujuh kandidat gen seleksi positif diidentifikasi dalam jalur melanogenesis ( POMC , MAP2K2 , FZD5 , CREB3L3 , WNT5B , FZD10 , dan CALM1 ). Di antara gen-gen ini, tiga, yaitu WNT5B , FZD5 , dan FZD10 , dikaitkan dengan jalur pensinyalan Wnt, yang juga mencakup gen-gen seperti PPP3CA , CCND2 , CTNND2 , LGR6 , SIRT1 , WISP1 , dan LGR4 . Jalur pensinyalan Wnt, yang diketahui menginduksi stratifikasi dan regenerasi epidermis (Zhu et al., 2014 ), menunjukkan pengayaan signifikan pada unta AP, yang mungkin terkait dengan adaptasinya terhadap lingkungan gurun (Al Abri et al., 2023 ; Bahbahani et al., 2024 ). Secara khusus, gen FZD10 tampaknya memiliki hubungan dengan pembentukan melanin (Nie et al., 2022 ), yang menunjukkan peran potensialnya dalam mengatur pigmentasi, aspek penting perlindungan terhadap radiasi matahari. Gen lain yang menarik adalah BCO2 , yang telah dikaitkan dengan proses pigmentasi (Wu et al., 2021 ). Kehadirannya dalam populasi AP menggarisbawahi signifikansinya dalam adaptasi kulit dan bulu terhadap radiasi matahari yang intens di wilayah tersebut. Studi kami juga mengidentifikasi dalam populasi AP kandidat tanda tangan seleksi positif di suatu wilayah termasuk SIRT1 . Pengamatan ini menunjukkan bahwa sifat genetik populasi ini responsif terhadap radiasi matahari (Tian et al., 2020 ). SIRT1 dikenal karena perannya dalam berbagai proses seluler, termasuk perbaikan DNA dan pengaturan respons stres oksidatif, yang sangat penting untuk mengurangi kerusakan akibat paparan radiasi matahari. Temuan ini mendukung bahwa populasi ini beradaptasi untuk meminimalkan dampak negatif radiasi matahari.
KESIMPULAN
Studi ini mendokumentasikan keragaman genetik, struktur populasi, dan adaptasi populasi ayam lokal AP terhadap lingkungan iklim yang keras dalam konteks nasional dan global. Hasil kami menunjukkan pengelompokan semua populasi menurut wilayah asal geografisnya, dengan diferensiasi genetik minimal yang diamati dalam populasi tersebut. Sejauh pengetahuan kami, ini adalah analisis sekuens genom utuh skala besar pertama yang menargetkan ayam lokal dari AP, yang meliputi wilayah timur Arab Saudi dan Oman.
Dengan mempertimbangkan interaksi yang terdokumentasi dengan baik antara evolusi spesies dan faktor lingkungan, studi ini juga mengeksplorasi tanda tangan genetik seleksi positif dalam populasi yang beradaptasi dengan rentang suhu yang berbeda di berbagai lokasi geografis. Investigasi kami juga melibatkan identifikasi sejumlah gen yang terkait dengan adaptasi lingkungan dalam setiap populasi, serta diferensiasi profil gen antara populasi-populasi ini. Studi ini mengungkap serangkaian gen yang terkait dengan berbagai fungsi, termasuk pengaturan metabolisme dan energi ( SUGCT , HECW1 , MMADHC ), pengembangan sistem kardiovaskular dan angiogenesis ( RYR2 , LDB2 , SOX5 , FHOD3 ), apoptosis dalam sel ( APP , SRBD1 , NTN1 , PUF60 , SLC26A8 , DAP , SUGCT ), perlindungan terhadap radiasi matahari ( FZD10 , BCO2 , WNT5B , COL6A2 , SIRT1 ), dan pertumbuhan ( NELL1 ). Gen-gen ini mungkin berperan dalam adaptasi terhadap stres termal, berpotensi dengan alel atau haplotipe yang berbeda yang berkontribusi terhadap adaptasi terhadap stres dingin atau panas. Namun, penting untuk menekankan bahwa penyelidikan lebih lanjut diperlukan untuk menyelidiki lebih dalam temuan-temuan ini. Namun, hasil kami memberikan dukungan yang berharga bagi keberadaan adaptasi genetik di antara populasi yang diteliti yang beradaptasi dengan kondisi lingkungan yang berbeda.



Tinggalkan Balasan