
Abstrak
Bahasa Indonesia: Untuk menyelidiki keragaman genetik pada jintan hitam ( Nigella sativa L.), total 64 genotipe diuji dalam desain kisi sederhana 8 × 8 di Pusat Penelitian Pertanian Sirinika selama dua tahun berturut-turut selama 2022/2023 dan 2023/2024. Analisis varians mengungkapkan adanya perbedaan nyata di antara genotipe untuk semua sifat kuantitatif kecuali panjang dan diameter kapsul. Variasi genotipe untuk hasil biji berada di antara 0,66 dan 1,69 t ha −1 dengan rata-rata 1,02 t ha −1 . Koefisien variasi genotipik (GCV) dan koefisien variasi fenotipik (PCV) untuk 16 sifat diperkirakan dalam kisaran antara 3,81% dan 49,84% dan 6,50% dan 50,90%, berturut-turut. H 2 dan kemajuan sebagai persentase rata-rata (GAM) berkisar dari 19,18 hingga 98,25 dan 4,67% dan 100,70%, masing-masing. Estimasi GCV, PCV, H 2 , dan GAM yang tinggi hingga sedang diamati untuk semua parameter kecuali hari pembentukan kuncup, hari hingga matang, panjang kapsul, dan diameter kapsul. Hasil biji menunjukkan korelasi positif dan signifikan dengan hampir semua parameter. Lima PCA pertama menyumbang 72% dari total variabilitas, di mana PCA1 berkontribusi 32% dan PCA2, PCA3, PCA4, dan PCA5 masing-masing menunjukkan kontribusi 13%, 12%, 8%, dan 7% terhadap total variabilitas. Data kualitatif menunjukkan variabilitas tinggi kecuali warna bunga, tipe pertumbuhan tanaman, dan rambut batang. Analisis klaster untuk sifat kualitatif diklasifikasikan menjadi lima kelompok. Hasil penelitian dengan jelas menunjukkan bahwa ada keragaman yang cukup di antara genotipe, yang dapat dimanfaatkan untuk program perbaikan jintan hitam di masa mendatang.
Ringkasan Bahasa Sederhana
Uji lapangan dilakukan untuk menyelidiki keragaman genetik pada jintan hitam ( Nigella sativa L.) menggunakan 64 genotipe yang diuji dalam desain kisi sederhana 8 × 8 di Pusat Penelitian Pertanian Sirinka yang terletak di Ethiopia Timur Laut selama dua tahun berturut-turut selama 2023 dan 2024. Analisis varians mengungkapkan adanya perbedaan nyata di antara genotipe untuk semua sifat kuantitatif kecuali panjang dan diameter kapsul. Variasi genotipe untuk hasil biji berada di antara 0,66 dan 1,69 t ha −1 dengan rata-rata 1,02 t ha −1 . Estimasi tinggi hingga sedang dari koefisien variasi genotipe, koefisien variasi fenotipik, heritabilitas, dan kemajuan sebagai persentase rata-rata diamati untuk semua parameter. Lima sumbu komponen utama pertama/PCA/ menyumbang 72% dari total variabilitas, di mana PCA1 menyumbang 32% dan PCA2, PCA3, dan PCA4 masing-masing menunjukkan sumbangan 13%, 12%, 8%, dan 7% terhadap total variabilitas. Data kualitatif menunjukkan variabilitas yang tinggi dalam genotipe.
Singkatan
Bahasa Inggris
kemajuan genetik
GAM
kemajuan genetik sebagai persentase rata-rata
GCV
koefisien variasi genotipe
PCA
sumbu komponen utama
PCV
koefisien variasi fenotipik
SARC
Pusat Penelitian Pertanian Sirinika
Bahasa Indonesia: SAS
sistem analisis statistik
Universitas Gadjah Mada (UPGMA)
metode pengelompokan berpasangan tanpa bobot dengan rata-rata aritmatika
1. PENDAHULUAN
Tanaman obat telah digunakan oleh manusia sejak lama; salah satunya adalah jintan hitam ( Nigella sativa L.) yang telah digunakan untuk perawatan kesehatan sejak peradaban manusia. Jintan hitam juga dikenal sebagai biji hitam, bunga adas, ketumbar Romawi, dan biji bawang liar (Kulloli, 2016 ). Jintan hitam merupakan anggota Ranunculaceae (Weiss, 2002 ). Jintan hitam diyakini berasal dari wilayah Mediterania. Jintan hitam dibudidayakan secara luas di Afrika Utara, Eropa Selatan, Timur Tengah, Tiongkok, Jepang, Turki, Iran, dan anak benua India (Bhutia et al., 2015 ; Rana et al., 2012 ).
Mayoritas pasokan komersial global benih ini diperoleh dari budidaya di India (Dessie et al., 2020 ). Estimasi produksi dunia awal adalah 600.000 ton, dengan pangsa 86% di India, diikuti oleh Iran (4%), Suriah (3%), dan Turki (2%). Sisanya 5% berasal dari berbagai negara seperti Mesir, Pakistan, dan Afghanistan (CBI, 2023 ). Sebaliknya, 10 negara teratas yang memiliki nilai ekspor jintan hitam tertinggi secara berurutan adalah Malaysia, Indonesia, Tiongkok, Belanda, Jerman, Swedia, India, Amerika Serikat, Rusia, dan Spanyol (Tridge, 2023 ).
Di Ethiopia, jintan hitam merupakan salah satu tanaman rempah terpenting dan terutama diproduksi untuk memberi rasa pada makanan, menyiapkan minyak untuk parfum dan obat-obatan, sebagai sumber pendapatan, untuk diversifikasi tanaman, dan untuk tujuan ekspor (Desalegn & Wubshet, 2018 ; Wubshet & Desalegn, 2019 ). Ini merupakan tanaman terpenting kedua, yang diekspor ke pasar internasional setelah jahe (Teshome & Anshiso, 2019 ). Ethiopia adalah negara dengan zona agroekologi yang berbeda dan menguntungkan untuk produksi berbagai rempah-rempah, sayuran, dan tanaman (Dessie, Koye, et al., 2019 ; Dessie, Abtew, et al., 2019 ). Banyak varietas rempah-rempah, terutama jintan hitam, jintan putih, lada, kunyit, fenugreek, bawang putih, ketumbar, jahe, kapulaga, dan kemangi ditanam di Ethiopia untuk konsumsi dan tujuan komersial (Tesfa et al., 2017 ).
Meskipun kondisi lingkungan negara tersebut menguntungkan untuk produksi jintan hitam dan varietasnya yaitu, Silingo, Kena, Gemechis, Darbera, Soressaa, Keneni, Dershaye, dan Eden yang direkomendasikan, produktivitas rata-rata nasional sangat rendah dan dilaporkan sebesar 0,64 t ha −1 , yang berada di bawah rata-rata global dan beberapa negara penghasil utama, termasuk India (2,20 t ha −1 ) (Organisasi Pangan dan Pertanian, 2023 ; Zigyalew, 2020 ). Kurangnya varietas unggul, masalah penanganan pascapanen, dan praktik pertanian yang tidak memadai merupakan faktor utama yang memengaruhi produksi dan produktivitasnya (Getachew & Beriso, 2020 ).
Penilaian keragaman genetik dapat dilakukan melalui penggunaan berbagai data seperti data morfologi dan agronomi, silsilah, biokimia, dan molekuler. Sifat morfologi dikendalikan secara genetik oleh jumlah gen dan ekspresinya bergantung pada faktor lingkungan dalam sebagian besar kasus (Abdi et al., 2002 ; Tuinstra et al., 1996 ; Van Beuningen & Busch, 1997 ).
Terdapat sifat morfologi kuantitatif dan kualitatif yang memainkan peran penting dalam klasifikasi individu. Sifat morfologi kualitatif mengalami pengaruh lingkungan yang relatif lebih sedikit dan memiliki kendali mono- atau oligogenik (Lima et al., 2017 ). Sifat-sifat tersebut tidak memiliki nilai kuantitatif dan mungkin biner atau multikategori. Pemanfaatan data morfologi kualitatif merupakan strategi yang bermanfaat dalam studi keragaman genetik tidak hanya karena merupakan aplikasi praktis dan berbiaya rendah, tanpa persyaratan peralatan yang canggih, tetapi juga diharapkan dapat memberikan informasi tambahan tentang unit hierarkis.
Variasi pada karakteristik yang sangat dapat diwariskan dan sifat kualitatif memberikan perkiraan keanekaragaman, sementara sifat kuantitatif dimanfaatkan untuk menentukan besarnya variasi genetik dalam plasma nutfah (Iqbal et al., 2010 ). Selain itu, evaluasi stok genetik yang tersedia untuk menilai variasi genetik untuk karakteristik yang penting secara ekonomi merupakan prasyarat untuk menggabungkan gen yang diinginkan dalam satu aksesi (Palevitch, 2004 ).
Sangat sedikit penelitian yang dilakukan untuk menentukan keragaman genetik pada sifat-sifat kuantitatif di lokasi tertentu selama 1 tahun di Ethiopia. Menurut Basazinew et al. ( 2023 ) dengan 64 aksesi di Debre-Zeit dan Kulumsa, ditemukan bahwa ada keragaman yang cukup di antara genotipe, yang dapat dimanfaatkan untuk program perbaikan jintan hitam di masa mendatang. Selain itu, Yewubdinber ( 2020 ) dan Yewubdinber et al. ( 2022 ) dengan 25 aksesi di Debre-Zeit melaporkan adanya variasi di antara genotipe dan menyarankan evaluasi lebih lanjut terhadap plasma nutfah jintan hitam di Ethiopia terhadap hasil biji. Namun, tidak ada penelitian keragaman yang dilakukan pada sifat-sifat kualitatif dan tidak ada cukup informasi berdasarkan sifat-sifat kuantitatif. Oleh karena itu, penelitian ini dilakukan untuk menyelidiki keragaman genotipe jintan hitam Ethiopia berdasarkan sifat-sifat kuantitatif dan kualitatif.
2 BAHAN DAN METODE
2.1 Lokasi tempat percobaan
Percobaan lapangan dilakukan untuk mengevaluasi keragaman genetik aksesi jintan hitam. Percobaan dilakukan di Pusat Penelitian Pertanian Sirinika (SARC), Ethiopia Timur Laut. SARC terletak pada 11°45′00″ Lintang Utara dan 39°36′36″ Bujur Timur dengan ketinggian 1850 m di atas permukaan laut. Curah hujan tahunan untuk tahun 2022/2023 dan 2023/2024 masing-masing adalah 101,64 dan 105,29 mm. Dengan cara yang sama, suhu minimum-maksimum untuk tahun 2022/2023 dan 2023/2024 masing-masing adalah 13,49/26,35 dan 13,72/26,59 0°C. Jenis tanah yang dominan di lokasi tersebut adalah lempung liat (SARC [Pusat Penelitian Pertanian Sirinika], 2023 ).
2.2 Bahan tanaman dan desain eksperimen
Enam puluh empat aksesi jintan hitam yang diperoleh dari Ethiopian Biodiversity Institute (EBI) digunakan dalam penelitian ini (Tabel S1 ). Saya menggunakan jumlah maksimum genotipe yang dikumpulkan dari daerah potensial pertumbuhan jintan hitam di Ethiopia oleh EBI. Bahan dievaluasi menggunakan desain kisi sederhana 8 × 8 pada musim tanam utama dari Juli hingga Januari tahun 2022/2023 dan 2023/2024. Ukuran plot untuk setiap unit percobaan adalah 1,2 × 2,1 m (empat baris). Total luas plot adalah 2,52 m 2 . Jarak antar tanaman, baris, plot, dan blok masing-masing adalah 0,15, 0,30, 0,50, dan 2,00 m. Penaburan dilakukan dengan pengeboran tangan dan ditutup sedikit dengan tanah (Habtewold et al., 2017 ). Pupuk dengan takaran 64,90 kg ha -1 N dan 37,70 kg ha -1 P 2 0 5 digunakan untuk memperlancar dan meningkatkan perkembangan akar serta hasil tanaman jintan hitam (Amdie et al., 2024 ). Penyiangan, penjarangan, dan perlindungan tanaman dilakukan setelah penanaman (Habtewold et al., 2017 ).
2.3 Data yang dikumpulkan
Data kualitatif dikumpulkan pada kerapatan cabang, pola percabangan, bulu kapsul, warna bunga, laju pembungaan, waktu pembungaan, ukuran folikel, daya tumbuh, luas daun, warna daun, warna batang utama, habitus pertumbuhan tanaman, tipe pertumbuhan tanaman, sistem perakaran, pematangan biji, elastisitas batang, bulu batang, bentuk batang pada penampang melintang, ketebalan batang, dan tingkat kepedasan kapsul. Selain itu, data kuantitatif dikumpulkan pada hari pembentukan tunas, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, kapsul per tanaman, biji per kapsul, diameter kapsul, panjang kapsul, tinggi tanaman, cabang primer per tanaman, cabang sekunder per tanaman, berat 1000 biji, hasil biji per tanaman, hasil biji per hektar, hasil biomassa per hektar, dan indeks panen. Daftar deskriptor untuk mengkarakterisasi jintan hitam sejauh ini belum tersedia. Oleh karena itu, deskriptor tanaman umum menggunakan Bioversity International ( 2007 ) digunakan untuk mencatat dan menganalisis karakter kualitatif (Tabel S2 ). Data karakteristik kuantitatif diperoleh secara plot dan diukur dari 10 tanaman representatif, kemudian diperoleh hasil rata-ratanya.
2.4 Analisis data
2.4.1 Analisis varians dan perbandingan mean
Semua data kuantitatif yang dihasilkan dianalisis menggunakan perangkat lunak Statistical Analysis System (SAS) (SAS, 2019 ). Estimasi komponen varians dihitung dengan menyamakan kuadrat rata-rata yang diamati dari analisis varians (ANOVA) dengan kuadrat rata-rata yang diharapkan. Sebelum melakukan ANOVA gabungan selama bertahun-tahun, data setiap tahun dikenakan ANOVA dan uji normalitas menggunakan uji Shapiro–Wilk W. Uji Bartlett digunakan untuk menentukan homogenitas varians galat dari eksperimen tahun individual, dan kemudian ANOVA gabungan selama bertahun-tahun dilakukan setelah mengonfirmasi homogenitas varians. ANOVA gabungan dibuat menggunakan prosedur Generalized Linear Model (GLM) untuk desain kisi sederhana sebagai berikut:
![]()
di mana X ijkl adalah nilai teramati dari genotipe i pada blok l , replikasi k tahun j , μ adalah rata-rata agung, G i adalah pengaruh genotipe i , Y j adalah pengaruh tahun j , GY ij adalah pengaruh interaksi genotipe i dengan tahun j , R k adalah pengaruh replikasi k , B l ( k ) adalah pengaruh blok l pada replikasi k , dan ε ijkl adalah pengaruh galat (residual) genotipe i pada blok l , replikasi k tahun j .
2.4.2 Variabilitas fenotipik dan genotipik
Varians fenotipik ( σ 2 p) dan genotipik ( σ 2 g) serta koefisien variasi fenotipik (PCV) dan koefisien variasi genotipik (GCV) yang sesuai untuk setiap sifat dihitung mengikuti metode yang dijelaskan oleh Burton dan De Vane ( 1953 ).
Varians genotipe diperkirakan sebagai berikut:
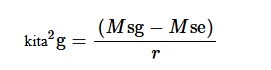
di mana σ 2 g merupakan varians genotipe, r merupakan jumlah replikasi, M sg merupakan kuadrat rata-rata genotipe, dan M se merupakan kuadrat rata-rata kesalahan.
Varians fenotipik diperkirakan mengikuti varians genotipik yang diperkirakan (Burton & De vane, 1953 ) sebagai berikut:
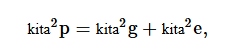
di mana σ 2 p merupakan varians fenotipik, σ 2 g merupakan varians genotipik, dan σ 2 e merupakan varians lingkungan.
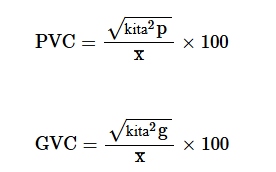
di mana PCV merupakan koefisien variasi fenotipik, GCV merupakan koefisien variasi genotipik, dan X merupakan rata-rata populasi dari sifat yang dievaluasi.
Nilai PCV dan GCV dikategorikan menjadi nilai rendah, sedang, dan tinggi seperti yang ditunjukkan oleh Sivasubramaniah dan Menon ( 1973 ) sebagai berikut: rendah = 0%–10%, sedang = 10%–20%, dan tinggi = >20%.
2.4.3 Heritabilitas dan kemajuan genetik (GA)
Nilai heritabilitas dalam arti luas diestimasi menggunakan rumus yang diadopsi oleh Falconer dan Mackay ( 1996 ) sebagai berikut:
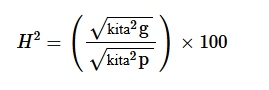
dimana H 2 adalah heritabilitas dalam arti luas, σ 2 p adalah varians fenotipik, dan σ 2 g adalah varians genotipik.
Persentase heritabilitas dikategorikan sebagai rendah, sedang, dan tinggi seperti yang disarankan oleh Robinson et al. ( 1955 ) dan Johnson et al. ( 1955 ): rendah = 0%–30%, sedang = 30%–60%, dan tinggi = >60%.
Diharapkan GA sedang dalam seleksi
GA dalam satuan absolut dan persentase rata-rata (GAM), dengan asumsi pemilihan 5% genotipe unggul, diperkirakan sesuai dengan metode yang diilustrasikan oleh Johnson et al. ( 1955 ) sebagai berikut:

di mana GAM merupakan kemajuan genetik sebagai persentase rata-rata, GA, dan x merupakan rata-rata populasi untuk sifat yang dievaluasi.
GA sebagai persentase rata-rata dikategorikan sebagai rendah, sedang, dan tinggi seperti yang disarankan oleh Johnson et al. ( 1955 ) sebagai berikut: rendah = 0%–10%, sedang = 10–20, dan tinggi = >20.
2.4.4 Analisis komponen utama
Analisis komponen utama (PCA) adalah salah satu teknik statistik multivariat, yang merupakan alat yang ampuh untuk menyelidiki dan meringkas tren yang mendasari dalam struktur data yang kompleks (Legendre & Legendre, 1998 ). Data distandarisasi untuk berarti nol dan varians satu sebelum menghitung analisis komponen utama. Analisis ini mencerminkan pentingnya kontributor terbesar terhadap variasi total pada setiap sumbu untuk diferensiasi (Sharma, 1998 ). Holland ( 2008 ) menyarankan bahwa kriteria standar mengizinkan pengabaian komponen yang variansnya dijelaskan <1 ketika matriks korelasi digunakan. Menurut Brejda et al. ( 2000 ), data dipertimbangkan dalam setiap komponen dengan nilai eigen >1, yang menentukan setidaknya 10% dari variasi. Nilai eigen yang lebih tinggi dianggap sebagai representasi terbaik dari atribut sistem dalam komponen utama.
2.4.5 Analisis data korelasi
Korelasi fenotipik antara dua sifat diperkirakan menggunakan rumus yang disarankan oleh Johnson et al. ( 1955 ) dan Singh dan Chaudhury ( 1985 ).
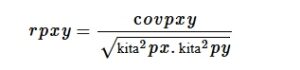
di mana rpxy adalah koefisien korelasi fenotipik antara karakter x dan y ; cov pxy adalah kovariansi fenotipik antara karakter x dan y σ 2 px adalah varians fenotipik untuk karakter x σ 2 py adalah varians fenotipik untuk karakter y.
Koefisien korelasi pada tingkat fenotipik diuji signifikansinya dengan membandingkan nilai koefisien korelasi dengan nilai r tabulasi pada derajat kebebasan g-2, di mana “g” adalah jumlah genotipe. Namun, koefisien korelasi pada tingkat genotipe diuji signifikansinya menggunakan rumus yang dijelaskan oleh Robertson ( 1959 ).
2.4.6 Analisis data kualitatif dan pengelompokan
Distribusi frekuensi morfologi diestimasi berdasarkan karakter yang diamati pada aksesi. Untuk menilai keragaman fenotip semua aksesi, indeks keragaman Shannon–Weaver ( H ′) dihitung menggunakan frekuensi fenotip. Shannon–Weaver diestimasi menggunakan rumus berikut (Shannon, 1949 ):
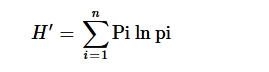
di mana pi adalah proporsi aksesi di kelas ke – i dari sifat tersebut terhadap jumlah total aksesi yang dikelompokkan dalam sifat tersebut, di mana “ n ” adalah jumlah kelas fenotip dalam sifat tersebut. Seperti yang diadopsi oleh Islam et al. ( 2019 ), H ′ diklasifikasikan sebagai rendah ( H ′ < 0,50), sedang ( H ′ = 0,50–0,75), dan tinggi ( H ′ ≥ 0,75). Analisis pengelompokan dilakukan berdasarkan metode pengelompokan berpasangan tak terbobot dengan rata-rata aritmatika (UPGMA). Jumlah klaster ditentukan menggunakan kriteria pengelompokan kubik seperti yang dijelaskan oleh Mohammadi dan Prasanna ( 2003 ). Jumlah klaster yang tepat ditentukan dari nilai pseudo-F dan pseudo-T2 menggunakan perangkat lunak SAS (SAS, 2019).
3 HASIL
3.1 ANOVA dan kinerja rata-rata
Karena setiap tahun ANOVA menunjukkan variansnya normal dan homogen, analisis gabungan dilakukan. ANOVA gabungan selama 2 tahun menunjukkan perlakuan menunjukkan efek yang signifikan ( p ≤ 0,01) kecuali untuk diameter kapsul dan diameter kapsul. Namun, efek interaksi tahun × genotipe tidak signifikan dalam semua sifat kuantitatif yang dipelajari (Tabel 1 ). Selain itu, sifat kualitatif menunjukkan perbedaan keragaman sedang hingga tinggi di antara 64 genotipe jintan hitam Ethiopia (Gambar S1 ).
| Bahasa Inggris | df | DBF | DCF | DF | DM | Bahasa Indonesia | NPB | Bahasa Inggris Raya | TWB | NCP | KL | CD | NSC | SIPP | SIPH | OLEHPH | HAI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Replikasi | 1 | 26.32** | 5.54* | 2,30 menit | 10.57** | 3,26 detik | 16.73** | 13.89** | 1,18 detik | 14.58** | 6.93** | 0,08 detik | 8.95** | 40.32** | 63.97** | 206.75** | 9.46** |
| Tahun | 1 | 8.46** | 73.35** | 10.75** | 90.55** | 12.64** | 8.17** | 45.94** | 54.44** | 65.24** | 54.06** | 1,72 detik | 4.60* | 8.16** | 3,29 detik | 1.05* | 1,44 detik |
| Memblokir | 7 | 0,82 detik | 5.97** | 2.16* | 5.10** | 0,53 detik | 0,67 detik | 1,27 detik | 1,75 detik | 2.36* | 2.15* | 1,25 detik | 1,95 detik | 0,95 detik | 0,81 detik | 3.46** | 1,33 detik |
| Blok (Replikasi) | 7 | 1,56 detik | 2,08 detik | 2.76* | 3.13** | 0,87 detik | 0,22 detik | 1,11 detik | 2.81** | 3.29** | 1,71 detik | 1,98 detik | 2,08 detik | 2,13 detik | 0,97 detik | 1,66 detik | 1,70 detik |
| Perlakuan | 63 | 6.01** | 41.13** | Tanggal 13.03** | 5.86** | 2.26** | 2.08** | 4.70** | Jam 7.30** | 24.81** | 0,79 detik | 1,24 detik | 3.63** | 57.28** | 10.27** | pukul 30.30** | 24.38** |
| Tahun × pengobatan | 63 | 1,2 detik | 1,15 detik | 1,44 detik | 0,41 detik | 0,97 detik | 1,02 detik | 1,36 detik | 1,29 detik | 0,74 detik | 1,15 detik | 1,2 detik | 0,40 detik | 0,99 detik | 0,46 detik | 0,40 detik | 0,60 detik |
| Sisa | 113 | 14.63 | 6.23 | 9.31 | Tanggal 22.18 | 30.54 | 1.24 | 4.54 | 0,05 | 7.29 | 0,02 | 0,01 | 106.52 | 0.12 | 0,02 | 0.34 | 6.78 |
| Riwayat Hidup | 6.21 | 3.05 | 4.38 | 3.80 | pukul 14.55 | 19.71 | 13.88 | 8.46 | tanggal 12.22 | Tanggal 12.13 | 11.97 | Tanggal 12.03 | 6.10 | 12.73 | 9.41 | 14.58 | |
* dan ** signifikan pada 0,05 dan sangat signifikan pada 0,01. ns: tidak signifikan pada 0,05. Singkatan: BYPH, hasil biomassa per hektar (t ha −1 ); CD, diameter kapsul (cm); CL, panjang kapsul (cm); DBF, hari hingga pembentukan kuncup; DCF, hari hingga pembentukan kapsul; DF, hari hingga 50% pembungaan; DM, hari hingga matang; HI, indeks panen (%); NCP, kapsul per tanaman; NPB, cabang primer per tanaman; NSB, cabang sekunder per tanaman; NSC, biji per kapsul; PH, tinggi tanaman (cm), SV, sumber variasi; SYPH, hasil biji per hektar (t ha −1 ); SYPP, hasil biji per tanaman (g); TSW, berat 1000 biji (g).
3.1.1 Rata-rata kinerja atribut fenologi
Berdasarkan hasil yang disajikan pada Tabel S3 , terdapat perbedaan yang sangat nyata ( p ≤ 0,01) antar genotipe untuk hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, tinggi tanaman, cabang primer per tanaman, cabang sekunder per tanaman, dan berat 1000 biji dengan rentang yang lebih lebar antara nilai minimum dan maksimum. Performa hari pembentukan kuncup berkisar antara 53,75 hingga 80,25 hari, dengan nilai rata-rata 61,59 hari. Rata-rata hari pembentukan kuncup tertinggi diperoleh dari genotipe 23.704, 23.705, dan 23.706, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 28.585, 28.592, 28.593, 30.785, dan 19.935 (Tabel S3 ). Hasil ini menunjukkan bahwa jumlah hari tertinggi dan terendah menyiratkan keterlambatan dan ketepatan waktu genotipe untuk mencapai pembentukan kuncup, masing-masing. Hari-hari untuk kinerja pembentukan kapsul berkisar antara 69,25 hingga 107,75 hari, dengan nilai rata-rata 81,84 hari. Hari-hari rata-rata tertinggi untuk pembentukan kapsul diperoleh dari genotipe 23.704, 23.705, dan 23.706, sedangkan nilai rata-rata terendah diperoleh dari genotipe 28.591, 28.588, 30.787, 28.585, dan 28.595 (Tabel S3 ). Hasil ini menunjukkan jumlah hari tertinggi dan terendah menyiratkan keterlambatan dan ketepatan waktu genotipe untuk mencapai pembentukan kapsul, masing-masing. Hari-hari untuk kinerja pembungaan 50% berkisar antara 60,25 hingga 91,50 hari, dengan nilai rata-rata 69,59 hari. Rata-rata hari pembungaan 50% tertinggi diperoleh pada genotipe 23.706, sedangkan rata-rata terendah diperoleh secara merata pada lima genotipe yaitu 28.585, 30.788, 28.594, 28.593, dan 30.790 (Tabel 1). S3) .). Hasil ini menunjukkan jumlah hari tertinggi dan terendah menyiratkan keterlambatan dan ketepatan waktu genotipe untuk mencapai 50% pembungaan, berturut-turut. Kinerja hari hingga kematangan berkisar antara 114,00 hingga 149,75 hari, dengan nilai rata-rata 123,96 hari. Rata-rata hari hingga kematangan tertinggi diperoleh dari genotipe 23.704 dan 23.706, sedangkan nilai rata-rata terendah diperoleh secara merata dari tiga genotipe, yaitu 28.591, 19.935, dan 30.790. Hasil ini menunjukkan jumlah hari tertinggi dan terendah menyiratkan keterlambatan dan ketepatan waktu genotipe untuk mencapai kematangan, berturut-turut. Dengan demikian, genotipe 23.704 dan 23.706 matang terlambat, sedangkan genotipe 28.591, 19.935, dan 30.790 matang lebih awal. Jumlah cabang primer per kinerja tanaman berkisar antara 3,75 sampai 9,00, dengan nilai rata-rata 5,66. Jumlah cabang primer tertinggi diperoleh dari genotipe 23.704 dan 23.705, sedangkan nilai rata-rata terendah diperoleh secara merata dari tiga genotipe, yaitu 20.798, 20.790, dan 27.628. Jumlah cabang sekunder per kinerja tanaman berkisar antara 9,50 sampai 21,50, dengan nilai rata-rata 15,35. Jumlah cabang sekunder tertinggi diperoleh dari genotipe 31.106, 23.705, 9.067, 19.926, 23.706, dan 9.068, sedangkan nilai rata-rata terendah diperoleh secara merata dari tiga genotipe, yaitu 20.794, 31.112, dan 20.793. Performa tinggi tanaman berkisar antara 28,75 sampai 49,59 cm, dengan nilai rata-rata 37,99 cm. Rata-rata tinggi tanaman tertinggi diperoleh dari genotipe 20.941, 23.705, 23.704, dan 23.706, diikuti oleh genotipe 19.926, 31.108, 9068, 31.109, 27.627, 31.114, 19.931, dan 31.107, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 28.595, 30.789, 30.791, 30.789, dan 20.790 (Tabel S3 ). Performa berat seribu biji berkisar antara 2,18 sampai 4,06 g, dengan nilai rata-rata 2,64 g. Rata-rata bobot 1000 biji tertinggi diperoleh pada genotipe 31.106, disusul oleh genotipe 23.705, 23.704, 30.790, dan 19.935, sedangkan rata-rata terendah diperoleh secara merata pada tiga genotipe, yaitu 19.926, 27.626, dan 19.932 (Tabel S3 ).
3.1.2 Rata-rata kinerja pertumbuhan, hasil, dan atribut terkait hasil
Ada perbedaan yang signifikan antar genotipe untuk kapsul per tanaman, biji per kapsul, hasil biji per tanaman, hasil biji per hektar, hasil biomassa per hektar, dan indeks panen dengan rentang yang lebih lebar antara nilai minimum dan maksimum, tetapi genotipe tidak menunjukkan perbedaan yang signifikan dengan panjang dan diameter kapsul (Tabel S4 ). Jumlah kapsul per kinerja tanaman berkisar antara 11,75 hingga 43,25, dengan nilai rata-rata 22,11. Jumlah rata-rata kapsul tertinggi diperoleh dari genotipe 31.109, 31.114, 31.121, dan 30.786, sedangkan nilai rata-rata terkecil diperoleh secara merata dari lima genotipe, yaitu 30.793, 28.595, dan 30.790 (Tabel S4 ). Jumlah biji per kapsul berkisar antara 63,50 sampai 122,75, dengan nilai rata-rata 85,83. Jumlah biji rata-rata tertinggi diperoleh dari genotipe 9068 dan 19.926, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 20.797, 20.790, dan 28.588 (Tabel S4 ). Kinerja hasil biji per tanaman berkisar antara 3,37 sampai 9,15 g, dengan nilai rata-rata 5,60 g. Hasil biji rata-rata tertinggi diperoleh dari genotipe 31.121, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 25.888, 30.793, dan 27.629 (Tabel S4 ). Kinerja hasil biji berkisar antara 0,66 hingga 1,69 t ha −1 , dengan rata-rata 1,02 t ha −1 . Rata-rata hasil biji per hektar tertinggi diperoleh dari genotipe 20.942, 31.114, dan 9068, sedangkan nilai rata-rata terendah diperoleh dari 19.931, 27.629, dan 30.792 (Tabel S4 ). Kinerja hasil biomassa per hektar berkisar antara 2,78 hingga 9,29 t ha −1 , dengan nilai rata-rata 6,23 t ha −1 . Rata-rata hasil biomassa per hektar tertinggi diperoleh dari genotipe 31.121, 31.112, 31.109, 27.627, dan 9.068, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 28.586, 27.626, dan 20.792 (Tabel S4 ). Kinerja indeks panen berkisar antara 10,43% hingga 43,09%, dengan nilai rata-rata 17,85%. Rata-rata indeks panen tertinggi diperoleh dari genotipe 27.626 dan 20.792, sedangkan nilai rata-rata terendah diperoleh secara merata dari lima genotipe, yaitu 30.792, 31.106, 27.625, dan 19.931 (Tabel S4 ).
3.2 Estimasi fenotipik dan GCV
Perkiraan varians fenotipik ( σ 2 p) dan genotipik ( σ 2 g), dan PCV dan GCV diberikan dalam Tabel 2. GCV dan PCV berkisar dari 3,81% hingga 49,84% dan 6,50% hingga 50,90%, masing-masing, untuk 16 sifat dari 64 genotipe jintan hitam. Hasil biji per tanaman, hasil biji per hektar, hasil biomassa per hektar, dan indeks panen memiliki nilai GCV dan PCV yang tinggi, sementara hari pembentukan kapsul, hari hingga 50% pembungaan, tinggi tanaman, berat 1000 biji, dan biji per kapsul memiliki rentang GCV dan PCV sedang dari 10,75% hingga 16,21%. Selain itu, hari hingga matang dan panjang kapsul memiliki nilai GCV dan PCV yang rendah. Estimasi PCV sedang untuk hari pembentukan tunas (10,76%) dan diameter kapsul (12,31), sedangkan PCV tinggi untuk cabang primer (20,07%) dan sekunder (21,28). Estimasi GCV <10% untuk hari pembentukan tunas dan diameter kapsul, sedangkan GCV sedang untuk cabang primer (14,46%) dan sekunder (18,88%) (Tabel 2 ).
| Sifat-sifat | Jangkauan | Rata-rata ± SE | σ 2 gram | σ 2 p | H2O | GCV | PCV | Bahasa Inggris | GAM |
|---|---|---|---|---|---|---|---|---|---|
| DBF | 53,75-80,25 | 61,59 ±2,71 | 36.63 | 43.95 | 83.35 | 9.83 | 10.76 | pukul 11.40 | 18.51 |
| DCF | 69,25-107,75 | 81,84 ±1,76 | 124.97 | 128.08 | 97.57 | 13.66 | 13.83 | 22.78 | 27.84 |
| DF | 60,25-91,50 | 69,59 ±2,16 | 55.95 | 60.60 | 92.32 | 10.75 | Tanggal 11.19 | 14.83 | Tanggal 21.31 |
| DM | 114.00-149.75 | 123,96 ±3,33 | 53.91 | 65.00 | 82.94 | 5.92 | 6.50 | 13.79 | Tanggal 11.13 |
| Bahasa Indonesia | 28.75-49.585 | 37,99 ±3,91 | tanggal 19.19 | jam 34.45 | 55.68 | 11.53 | pukul 15.45 | 6.74 | 17.75 |
| NPB | 3,75-9,00 | 5,66 ±0,79 | 0.67 | 1.29 | 51.86 | 14.46 | tanggal 20.07 | 1.22 | 21.47 |
| Bahasa Inggris Raya | Jam 9.50-21.50 | 15,35 ±1,51 | 8.40 | 10.67 | 78.74 | 18.88 | Tanggal 21.28 | 5.31 | 34.57 |
| TWB | 2.18-4.06 | 2,64 ±0,14 | 0.16 | 0.18 | 86.29 | tanggal 15.18 | Tanggal 16.10 | 0.76 | 28.66 |
| NCP | 11.75-43.25 | 22,11 ±1,91 | 86.82 | 90.46 | 95.97 | 42.15 | 43.03 | 18.83 | 85.18 |
| KL | 1.05-1.31 | 1,17 ±0,10 | 0.00 | 0,01 | 26.60 | 3.81 | 8.51 | 0,05 | 4.67 |
| CD | 0,68-0,94 | 0,81 ±0,0 | 0.00 | 0,01 | tanggal 19.18 | 3.89 | Tanggal 12.31 | 0,04 | 4.87 |
| NSC | 63,50-122,75 | 85,83 ±7,30 | 140.19 | 193.45 | 72.47 | 13.80 | tanggal 16.21 | 20.79 | tanggal 24.23 |
| SIPP | 3.37-9.15 | 5,6 ±0,24 | 3.29 | 3.35 | 98.25 | 32.37 | 32.66 | 3.71 | 66.21 |
| SIPH | 0,66-1,69 | 1,02 ±0,10 | 0,08 | 0,09 | 90.26 | 27.80 | 29.48 | 0.56 | 54.90 |
| OLEHPH | 2.78-9.29 | 6,23 ±0,41 | 5.04 | 5.21 | 96.70 | 36.02 | 36.63 | 4.55 | 73.07 |
| HAI | 10.43-43.09 | 17,85 ±1,84 | 79.18 | 82.57 | 95.90 | 49.84 | 50.90 | 17.98 | 100.70 |
Singkatan: BYPH, hasil biomassa per hektar (t ha −1 ); CD, diameter kapsul (cm); CL, panjang kapsul (cm); DBF, hari hingga pembentukan kuncup; DCF, hari hingga pembentukan kapsul; DF, hari hingga 50% pembungaan; DM, hari hingga kematangan; GA, kemajuan genetik; GAM, kemajuan genetik sebagai persentase rata-rata; GCV, koefisien varians genetik; H 2 , heritabilitas dalam arti luas; HI, indeks panen (%); NCP, kapsul per tanaman; NPB, cabang primer per tanaman; NSB, cabang sekunder per tanaman; NSC, biji per kapsul; PCV, koefisien varians fenotipik; PH, tinggi tanaman (cm); SYPH, hasil biji per hektar (t ha −1 ); SYPP, hasil biji per tanaman (g); TSW, berat 1000 biji (g); σ 2 g, varians genotipik; σ 2 p, varians fenotipik.
3.3 Estimasi heritabilitas dan GA
Heritabilitas arti luas ( H 2 ) yang diestimasi untuk 16 sifat dari 64 genotipe jintan hitam ditemukan dalam kisaran 19,18% (diameter kapsul) hingga 98,25% (untuk hasil biji per tanaman), sementara GA sebagai persentase rata-rata berkisar antara 4,67% (untuk diameter kapsul) hingga 100,70% (untuk indeks panen) (Tabel 2 ). Dengan demikian, estimasi H 2 dan GAM keduanya tinggi untuk hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, jumlah cabang sekunder, berat 1000 biji, kapsul per tanaman, biji per kapsul, hasil biji per tanaman, hasil biji per hektar, hasil biomassa per hektar, dan indeks panen. Estimasi H 2 yang tinggi ditambah dengan nilai GAM sedang untuk hari hingga matang. Estimasi H 2 sedang ditambah dengan nilai GAM yang tinggi untuk tinggi tanaman dan cabang primer, sedangkan nilai rendah diestimasi baik untuk H 2 maupun GAM untuk panjang kapsul dan diameter kapsul.
3.4 Analisis komponen utama
Hasil analisis komponen utama dari 16 sifat kuantitatif disajikan dalam Tabel 3. Empat sumbu komponen utama pertama (PCA1 hingga PCA5) menjelaskan persentase bervariasi dari total varians yang berkisar dari 7% hingga 32% dan menjelaskan 72% dari total variasi dengan nilai eigen berkisar dari 1,222 hingga 5,50. PCA1 berkontribusi 32% terhadap total variabilitas di antara genotipe, sementara PCA2, PCA3, PCA4, dan PCA5 masing-masing menunjukkan kontribusi 13%, 12%, 8%, dan 7% terhadap total variabilitas. Hari pembentukan kapsul, tinggi tanaman, hari pembentukan kuncup, hari hingga 50% pembungaan, dan hari hingga kematangan memiliki kontribusi lebih tinggi terhadap PCA1. Indeks panen, diikuti oleh hasil biomassa per hektar dan berat 1000 biji, memiliki kontribusi lebih besar terhadap PCA2. Hasil biji per tanaman memiliki kontribusi yang relatif lebih besar terhadap total varians PCA3, diikuti oleh panjang kapsul dan diameter kapsul. Indeks panen, berat 1000 biji, cabang primer, panjang kapsul, dan cabang sekunder memiliki kontribusi yang relatif lebih besar terhadap total varians PCA4. Terakhir, biji per kapsul, cabang sekunder, cabang primer, hasil biji per hektar, dan diameter kapsul memiliki kontribusi yang relatif lebih besar terhadap total varians PCA5.
| Variabel | Kartu PCI | PCI II | PCIII | PCIV | PCV |
|---|---|---|---|---|---|
| Nilai eigen | 5.5 | 2.2 | 1.96 | 1.33 | 1.22 |
| Perbedaan | 3.3 | 0.24 | 0.63 | 0.11 | 0.26 |
| Proporsi (variabilitas) | 0.32 | 0.13 | 0.12 | 0,08 | 0,07 |
| Kumulatif | 0.32 | 0,45 | 0.57 | 0,65 | 0.72 |
| DBF | 0.334 | -0,054 | 0,145 | -0,007 | -0,105 |
| DCF | 0,366 tahun | -0,148 | 0.162 | -0,167 | -0,048 |
| DF | 0.32 | -0,157 | 0.238 | -0,259 | 0,069 tahun |
| DM | 0.32 | -0,166 | 0,036 hari | -0,181 | -0,073 |
| Bahasa Indonesia | 0,336 tahun | -0,013 | 0,25 | 0.106 | -0,022 |
| NPB | 0,267 tahun | 0,094 tahun | 0,097 tahun | 0.354 | -0,329 |
| Bahasa Inggris Raya | 0.232 | 0,097 tahun | -0,007 | 0.34 | -0,365 |
| TWB | 0.127 | 0.328 | -0,26 | 0,374 tahun | -0,178 |
| NCP | 0.194 | -0,007 | -0,199 | 0,029 | 0,258 |
| KL | -0,013 | 0.254 | 0,379 tahun | 0,355 | 0.296 |
| CD | -0,158 | 0.299 | 0.347 | 0.189 | 0.302 |
| NSC | 0.13 | 0.153 | 0.189 | -0,074 | 0.394 |
| SIPP | 0.253 | 0.166 | -0,454 | -0,012 | 0.266 |
| SIPH | 0.227 | -0,258 | 0,029 | 0.234 | 0.321 |
| OLEHPH | 0.188 | 0.443 | 0.113 | -0,289 | 0,015 |
| HAI | -0,015 | -0,542 | -0,073 | 0.417 | 0.241 |
Singkatan: BYPH, hasil biomassa per hektar (t ha −1 ); CD, diameter kapsul (cm); CL, panjang kapsul (cm); DBF, hari hingga pembentukan kuncup; DCF, hari hingga pembentukan kapsul; DF, hari hingga 50% pembungaan; DM, hari hingga matang; HI, indeks panen (%); NCP, kapsul per tanaman; NPB, cabang primer per tanaman; NSB, cabang sekunder per tanaman; NSC, biji per kapsul; PCA1, sumbu komponen utama pertama; PCA2, sumbu komponen utama kedua; PCA3, sumbu komponen utama ketiga; PCA4, sumbu komponen utama keempat; PCA5, sumbu komponen utama kelima, dan angka dalam tanda kurung menunjukkan kontribusi sifat pada PCA; PH, tinggi tanaman (cm); SYPH, hasil biji per hektar (t ha −1 ); SYPP, hasil biji per tanaman (g); TSW, berat 1000 biji (g).
3.5 Analisis data korelasi
3.5.1 Koefisien korelasi hasil dengan sifat lainnya
Hasil koefisien korelasi disajikan dalam Tabel 4. Hasil biji memiliki koefisien korelasi positif dan signifikan dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, tinggi tanaman (cm), cabang primer per tanaman, dan kapsul per tanaman pada tingkat fenotipik. Hasil biji per hektar berkorelasi tinggi dan positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, tinggi tanaman, dan kapsul per tanaman. Hasil biji per hektar juga berkorelasi positif dan signifikan dengan hari hingga matang dan cabang primer per tanaman. Ia juga berkorelasi positif dengan panjang kapsul, biji per kapsul, hasil biji per tanaman, hasil biomassa per hektar, dan indeks panen tetapi berkorelasi negatif dengan cabang sekunder per tanaman, berat 1000 biji, dan diameter kapsul (Tabel 4 ).
| Sifat | DBF | DCF | DF | DM | Bahasa Indonesia | NPB | Bahasa Inggris Raya | TWB | NCP | KL | CD | NSC | SIPP | SIPH | OLEHPH | HAI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DBF | ||||||||||||||||
| DCF | 0,96** | |||||||||||||||
| DF | 0,94** | 0,94** | ||||||||||||||
| DM | 0,86** | 0,83** | 0,81** | |||||||||||||
| Bahasa Indonesia | 0,78** | 0,81** | 0,79** | 0,73** | ||||||||||||
| NPB | 0,38** | 0,40** | 0,30* | 0,37** | 0,44** | |||||||||||
| Bahasa Inggris Raya | 0.14 | 0.21 | 0.21 | 0.21 | 0,36** | 0,54** | ||||||||||
| TWB | 0,02 | 0,01 | -0,02 | 0.12 | -0,02 | 0.12 | 0,01 | |||||||||
| NCP | 0,47** | 0,49** | 0,48** | 0,44** | 0,49** | 0.21 | 0.11 | 0.12 | ||||||||
| KL | 0.17 | 0.18 | 0.12 | 0,04 | 0,38** | 0.24 | 0.12 | -0,04 | -0,003 | |||||||
| CD | -0,35** | -0,42** | -0,38** | -0,46** | -0,31* | -0,16 | -0,24 | -0,13 | -0,30* | 0,42** | ||||||
| NSC | 0,38** | 0,34** | 0,42** | 0,36** | 0,44** | 0,25* | 0,07 | 0,07 | 0,29* | 0.19 | -0,01 | |||||
| SIPP | 0,29* | 0,26* | 0,27* | 0,33** | 0,28* | 0.16 | 0,09 | 0,48** | 0,48** | -0,03 | -0,26* | 0,28* | ||||
| SIPH | 0,44** | 0,38** | 0,37** | 0,31* | 0,42** | 0,28* | -0,001 | -0,01 | 0,40** | 0,01 | -0,20 | 0.22 | 0.20 | |||
| OLEHPH | 0.22 | 0.23 | 0,26* | 0.23 | 0,31* | 0.23 | 0,25* | 0,26* | 0.24 | 0,26* | 0.10 | 0,36** | 0,34** | 0.19 | ||
| HAI | -0,31* | -0,32** | -0,34** | -0,34** | -0,25* | -0,005 | -0,074 | -0,15 | -0,21 | -0,04 | 0.18 | -0,19 | -0,17 | 0,15 | -0,42** |
* dan ** signifikan pada tingkat probabilitas 0,05, dan sangat signifikan pada tingkat probabilitas 0,01. Ns: tidak signifikan pada tingkat probabilitas 0,05, Singkatan: BYPH, hasil biomassa per hektar (t ha −1 ); CD, diameter kapsul (cm); CL, panjang kapsul (cm); DBF, hari hingga pembentukan kuncup; DCF, hari hingga pembentukan kapsul; DF, hari hingga 50% pembungaan; DM, hari hingga matang; HI, indeks panen (%); NCP, kapsul per tanaman; NPB, cabang primer per tanaman; NSB, cabang sekunder per tanaman; NSC, biji per kapsul; PH, tinggi tanaman (cm); SYPH, hasil biji per hektar (t ha −1 ); SYPP, hasil biji per tanaman (g); TSW, berat 1000 biji (g).
3.5.2 Koefisien korelasi antar sifat lainnya
Hari pembentukan kuncup berkorelasi tinggi dan positif dengan hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, tinggi tanaman, cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar tetapi berkorelasi tinggi dan negatif dengan diameter kapsul. Hari pembentukan kuncup juga berkorelasi positif dan signifikan dengan hasil biji per tanaman dan berkorelasi negatif dan signifikan dengan indeks panen. Ia juga berkorelasi positif dengan cabang sekunder per tanaman, berat 1000 biji, panjang kapsul, dan hasil biomassa per hektar (Tabel 4 ). Hari pembentukan kapsul berkorelasi tinggi dan positif dengan hari pembentukan kuncup, hari hingga 50% pembungaan, hari hingga kematangan, tinggi tanaman, cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar tetapi berkorelasi tinggi dan negatif dengan diameter kapsul dan indeks panen. Hari pembentukan kapsul juga berkorelasi positif dan signifikan dengan hasil biji per tanaman dan berkorelasi negatif dan sangat signifikan dengan diameter kapsul dan indeks panen. Itu juga memiliki korelasi positif dengan cabang sekunder per tanaman, berat 1000 biji, panjang kapsul, dan hasil biomassa per hektar (Tabel 4 ). Hari hingga 50% pembungaan sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga matang, tinggi tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar. Hari hingga 50% pembungaan, juga berkorelasi positif dan signifikan dengan cabang primer per tanaman, hasil biji per tanaman, dan hasil biomassa per hektar berkorelasi negatif dan sangat signifikan dengan diameter kapsul dan indeks panen. Itu juga memiliki korelasi positif dengan cabang sekunder per tanaman dan panjang kapsul tetapi korelasi negatif dengan berat 1000 biji (Tabel 4 ). Hari hingga matang sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, tinggi tanaman, cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per tanaman. Umur panen juga berkorelasi positif dan signifikan dengan hasil biji per hektar dan berkorelasi negatif dan sangat signifikan dengan diameter kapsul dan indeks panen. Korelasi positif juga terjadi pada cabang sekunder per tanaman, berat 1000 biji, panjang kapsul, dan hasil biomassa per hektar (Tabel 4) .). Tinggi tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, cabang primer per tanaman, cabang sekunder per tanaman, kapsul per tanaman, panjang kapsul, biji per kapsul, dan hasil biji per hektar. Tinggi tanaman juga berkorelasi positif dan signifikan dengan hasil biji per tanaman dan hasil biomassa per hektar dan berkorelasi negatif dan signifikan dengan diameter kapsul dan indeks panen. Itu juga memiliki korelasi negatif dengan berat 1000 biji (Tabel 4 ). Cabang primer per tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, tinggi tanaman, dan cabang sekunder per tanaman. Cabang primer per tanaman juga berkorelasi positif dan signifikan dengan biji per kapsul, hasil biji per tanaman, dan hasil biji per hektar. Itu juga berkorelasi positif dengan berat 1000 biji, kapsul per tanaman, panjang kapsul, hasil biji per tanaman, dan hasil biomassa per hektar tetapi berkorelasi negatif dengan diameter kapsul dan indeks panen (Tabel 4 ). Cabang sekunder per tanaman sangat berkorelasi dengan tinggi tanaman dan cabang primer per tanaman. Cabang sekunder per tanaman juga berkorelasi positif dan signifikan dengan hasil biomassa per hektar. Itu juga berkorelasi positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, berat 1000 biji, kapsul per tanaman, panjang kapsul, biji per kapsul, dan hasil biji per tanaman tetapi berkorelasi negatif dengan diameter kapsul, hasil biji per hektar, dan indeks panen (Tabel 4 ). Berat seribu biji sangat berkorelasi dengan hanya hasil biji per tanaman. Berat seribu biji juga berkorelasi positif dan signifikan dengan hasil biomassa per hektar. Ia juga berkorelasi positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari kematangan, cabang primer per tanaman, cabang sekunder per tanaman, kapsul per tanaman, dan biji per kapsul tetapi berkorelasi negatif dengan hari hingga 50% pembungaan, tinggi tanaman, panjang kapsul, diameter kapsul, hasil biji per hektar, dan indeks panen (Tabel 4 ). Kapsul per tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari kematangan, tinggi tanaman, hasil biji per tanaman, dan hasil biji per hektar. Kapsul per tanaman juga berkorelasi positif dan signifikan dengan biji per kapsul dan berkorelasi negatif dan signifikan dengan diameter kapsul. Ia juga berkorelasi positif dengan cabang primer per tanaman, cabang sekunder per tanaman, berat 1000 biji, dan hasil biomassa per hektar tetapi berkorelasi negatif dengan panjang kapsul dan indeks panen (Tabel 4) .). Panjang kapsul berkorelasi sangat positif dan positif dengan tinggi tanaman dan diameter kapsul. Panjang kapsul juga berkorelasi positif dan signifikan dengan hasil biomassa per hektar. Ia juga berkorelasi positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, cabang primer per tanaman, cabang sekunder per tanaman, biji per kapsul, dan hasil biji per hektar tetapi berkorelasi negatif dengan berat 1000 biji, kapsul per tanaman, hasil biji per tanaman, dan indeks panen (Tabel 4 ) . Diameter kapsul berkorelasi sangat positif dengan panjang kapsul tetapi berkorelasi sangat negatif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, dan hari hingga matang. Diameter kapsul juga berkorelasi negatif dan signifikan dengan tinggi tanaman, kapsul per tanaman, dan hasil biji per tanaman. Itu juga berkorelasi positif dengan hasil biomassa per hektar dan indeks panen tetapi berkorelasi negatif dengan cabang primer per tanaman, cabang sekunder per tanaman, berat 1000 biji, biji per kapsul, dan hasil biji per hektar (Tabel 4 ). Biji per kapsul berkorelasi tinggi dan positif dengan hari pembentukan tunas, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga matang, tinggi tanaman, dan hasil biomassa per hektar. Biji per kapsul juga berkorelasi positif dan signifikan dengan cabang primer per tanaman, kapsul per tanaman, dan hasil biji per tanaman. Itu juga berkorelasi positif dengan cabang sekunder per tanaman, berat 1000 biji, panjang kapsul, dan hasil biji per hektar, tetapi berkorelasi negatif dengan diameter kapsul dan indeks panen (Tabel 4 ). Hasil biji per tanaman berkorelasi tinggi dan positif dengan hari hingga matang, berat 1000 biji, kapsul per tanaman, dan hasil biomassa per hektar. Hasil biji per tanaman juga berkorelasi positif dan signifikan dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, tinggi tanaman, dan biji per kapsul tetapi berkorelasi negatif dan signifikan dengan diameter kapsul. Ia juga berkorelasi positif dengan cabang primer per tanaman, cabang sekunder per tanaman, dan hasil biji per hektar tetapi berkorelasi negatif dengan panjang kapsul dan indeks panen (Tabel 4 ). Hasil biomassa per hektar berkorelasi tinggi dan positif dengan biji per kapsul dan hasil biji per tanaman tetapi berkorelasi negatif tinggi dengan indeks panen. Hasil biomassa per hektar juga berkorelasi positif dan signifikan dengan hari hingga 50% pembungaan, tinggi tanaman, cabang sekunder per tanaman, berat 1000 biji, dan panjang kapsul. Ia juga berkorelasi positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga matang, cabang primer per tanaman, kapsul per tanaman, diameter kapsul, dan hasil biji per hektar (Tabel 4) .). Indeks panen berkorelasi sangat negatif dengan hari pembentukan kapsul, hari pembungaan 50%, hari kematangan, dan hasil biomassa per hektar. Indeks panen juga berkorelasi negatif dan signifikan dengan hari pembentukan tunas dan tinggi tanaman. Indeks ini juga berkorelasi positif dengan diameter kapsul dan hasil biji per hektar tetapi berkorelasi negatif dengan cabang primer per tanaman, cabang sekunder per tanaman, berat 1000 biji, kapsul per tanaman, panjang kapsul, biji per kapsul, dan hasil biji per tanaman (Tabel 4 ).
3.6 Analisis data kualitatif
3.6.1 Distribusi dalam karakter
Aksesi menunjukkan variasi tinggi hingga sedang pada karakter kualitatif (Tabel S5 ). Kepadatan Cabang: 39,06% aksesi diamati rendah, 29,69% menunjukkan rata-rata, dan 31,25% menunjukkan tinggi (Tabel S5 ). Demikian pula, distribusi pola percabangan diamati tidak bercabang (15%), percabangan basal (44,06%), dan percabangan atas (40,94%) di antara aksesi. Kerontokan rambut kapsul aksesi lebih bervariasi, dengan 39,06% memiliki kerontokan rambut sedang, 26,56% memiliki rambut kuat atau banyak, 18,75% memiliki rambut lemah atau jarang, dan 15,53% memiliki rambut gundul (tidak ada rambut) (Tabel S5 ). Beberapa karakter menunjukkan variabilitas sedang, seperti warna bunga (62,50% putih dan 37,50% biru pucat), tipe pertumbuhan tanaman (60,94% determinan dan 39,06% tak tentu), dan bentuk batang pada penampang melintang (54,69% bulat dan 45,31% persegi). Lebih lanjut, aksesi menunjukkan variasi luas pada karakteristik daun berikut: warna daun (48,44% hijau, 28,13% hijau dengan corak kekuningan, 14,06% hijau dengan corak biru keabu-abuan, dan 9,38% hijau dengan corak ungu), luas daun (51,56% lebar lebar, 29,69% lebar rata-rata, dan 18,75% sempit) (Tabel S5 ). Elastisitas batang adalah 43,75% rendah, 40,63%, 17,81% sedang, dan 7,81% tinggi. Dengan cara yang sama, bulu batang diamati lemah atau jarang (42,19%), gundul (tidak ada bulu) (31,25%), dan sedang (26,56%), sedangkan ketebalan batang diamati sangat tebal (48,44%), tebal (20,31%), sedang (20,31%), dan sempit (10,94%). Tingkat pembungaan adalah sebagai berikut: 46,88% aksesi diamati sedang, 28,13% diamati rendah, dan 25% diamati tinggi (Tabel S5 ). Dengan cara yang sama, waktu pembungaan diamati menjadi pembungaan tengah (51,56%), pembungaan awal (37,5%), dan pembungaan akhir (10,94%) di antara aksesi. Ukuran folikel aksesi lebih bervariasi, dengan 40,63% memiliki ukuran sedang, 26,56% memiliki besar, 23,44% memiliki kecil, dan 9,38% memiliki sangat kecil (Tabel S5 ). Daya tumbuhnya adalah 26,56% sangat kuat, 25% kuat, 25% sedang, 14,06% lemah, dan 9,38% sangat lemah. Dengan cara yang sama, warna batang utama adalah sebagai berikut: 43,75% hijau keunguan, 21,88% ungu, 18,75% kuning, dan 15,63% hijau. Demikian pula, kebiasaan tumbuh tanaman diamati bersujud (43,75%), tegak (tidak berbulu) (31,25%), dan semi tegak (25,00%), sedangkan sistem perakaran diamati berupa akar tunggang tipis dalam (46,88%), serabut dangkal (28,13%), dan akar tunggang tebal berbonggol (25,00%). Demikian pula, distribusi pematangan benih diamati berupa pematangan awal (48,44%), pematangan sedang (37,50%), dan pematangan akhir (14,06%) di antara aksesi (Tabel S5 ). Selain itu, untuk tingkat kepedasan kapsul, 48,44%, 37,50%, dan 14,06% biji masing-masing teruji ringan, sedang, dan pedas.
3.6.2 Indeks keanekaragaman Shannon–Weaver ( H ′)
Dalam aspek ini, nilai H ′ untuk sebagian besar karakter fenotipik yang diamati menunjukkan tingkat keragaman normal di antara aksesi jintan hitam yang diuji, yang berkisar dari 0,66 untuk warna bunga hingga 1,53 untuk daya tumbuh, dengan nilai rata-rata 1,07 (Tabel S5 ). Berdasarkan klasifikasi ini, tiga karakter dikategorikan sebagai keragaman sedang, seperti warna bunga, tipe pertumbuhan tanaman, dan rambut batang (Tabel S5 ). Sisanya dari 16 karakter yaitu kerapatan cabang, pola percabangan, rambut kapsul, laju pembungaan, waktu pembungaan, ukuran folikel, daya tumbuh, luas daun, warna daun, warna batang utama, habitus pertumbuhan tanaman, sistem perakaran, pematangan biji, ketebalan batang, rambut batang, elastisitas batang, dan kepedasan kapsul memiliki keragaman genetik yang tinggi.
3.6.3 Analisis klaster
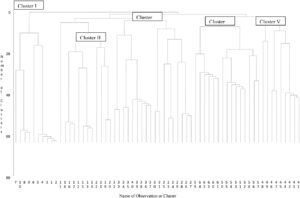
Matriks jarak Euclidean dari pasangan genotipe yang diestimasi dari sifat kuantitatif digunakan untuk menyusun dendrogram berdasarkan metode Unweighted Pair-group with Arithmetic Means (UPGMA). Sebanyak 64 genotipe jintan hitam dikelompokkan ke dalam lima klaster yang berbeda. Klaster III merupakan klaster terbesar dan terdiri dari 18 genotipe (28,13%) dari total genotipe. Klaster IV terdiri dari 15 (23,45%) dan tiga klaster lainnya (I, II, dan V) masing-masing terdiri dari 10 (15,61%), 12 (18,75%), dan sembilan (14,06%) (Tabel 5 ; Gambar 1 ).
| Gugus | Jumlah aksesi di setiap klaster | Daftar aksesi dalam klaster | % kontribusi |
|---|---|---|---|
| SAYA | 10 | G1, G2, G3, G4, G5, G6, G7, G8, G9, & G10 | 15.61 |
| II | 12 | G11, G12, G13, G14, G15, G16, G17, G18, G19, G20, G21, dan G22 | 18.75 |
| AKU AKU AKU | 18 | G23, G24, G25, G26, G27, G28, G29, G30, G31, G32, G33, G34, G35, G36, G37, G38, G39, & G40 | 28.13 |
| IV | 15 | G50, G51, G52, G53, G54, G55, G56, G57, G58, G59, G60, G61, G62, G63, dan G64 | pukul 23.45 |
| Bahasa Indonesia: V | 9 | G41, G42, G43, G44, G45, G46, G47, G48, dan G49 | tanggal 14.06 |
4 DISKUSI
4.1 Rata-rata kinerja atribut fenologi, pertumbuhan, hasil, dan terkait hasil
Dalam penelitian ini, hasil gabungan menunjukkan bahwa ada perbedaan yang sangat signifikan di antara genotipe untuk semua sifat kecuali panjang dan diameter kapsul dengan rentang yang lebih lebar antara nilai minimum dan maksimum. Hasil pendukung dilaporkan bahwa 36 genotipe jintan hitam memiliki 66–89 dan 117–150 hari hingga 50% pembungaan dan hari hingga matang, masing-masing, sebagaimana dievaluasi di Debre-Zeit (Tewodros et al., 2018 ). Demikian pula, 20 genotipe jintan hitam dievaluasi di tiga lokasi di dataran tinggi Bale (dievaluasi selama 2 tahun) dan dilaporkan memiliki 142–150 hari dengan rata-rata 145 hari hingga matang (Fufa, 2016 ). Jumlah cabang primer dan sekunder tertinggi tercatat untuk genotipe 30.763 (5,61) dan 30.754 (5,12), masing-masing, sementara 30.772 memiliki jumlah cabang primer (3,86) dan sekunder terendah (2,48) (Tewodros et al., 2018 ). Demikian pula, hasil serupa ditemukan dalam hasil biji untuk 10 ekotipe jintan hitam yang diuji oleh Golparvar et al. ( 2014 ). Laporan Gutiérrez-Prieto dan Kirici ( 2021 ) menunjukkan bahwa dalam kondisi Mediterania, siklus hidup tanaman berlangsung selama 225 hari. Periode vegetatif berlangsung selama 137 hari dan periode reproduksi 88 hari. Sementara itu, Haq et al. ( 2015 ) mengamati bahwa waktu yang dibutuhkan 50% tanaman untuk mencapai inisiasi kuncup bunga adalah 146 hari dan mendaftarkan kapsul pertama 78 hari setelah disemai. Dalam penelitian oleh Ürüşan ( 2016 ), tahap pembungaan dimulai sekitar 69,9 hari setelah penaburan dan berlangsung sekitar 43,9 hari. Tahap pengaturan kapsul untuk penelitian ini muncul 160 hari setelah penaburan, dan 100% kapsul telah matang pada hari ke-225. Selain itu, Asefa dan Beriso ( 2020 ) melaporkan bahwa hari hingga matang dan hari hingga pembungaan berkisar antara 144,83 hingga 128,00 dan 90,00 hingga 80,00, berturut-turut. Rata-rata total hasil biji tertinggi adalah 2,36 t ha −1 diikuti oleh 2,18 t ha −1 . Hal ini mungkin disebabkan oleh potensi distrik untuk produksi jintan hitam daripada kedua lingkungan sampel. Hasil serupa dilaporkan oleh Tewodros et al. ( 2018 ) pada tinggi tanaman jintan hitam, berat 1000 biji, hasil biji per tanaman, dan hasil biji per plot. Selain itu, Ermias et al. ( 2015 ) melaporkan tinggi tanaman berkisar antara 45,88 hingga 52,20 dengan tinggi tanaman rata-rata 49,43 cm di satu lokasi dan 39,12–51,36 dengan tinggi tanaman rata-rata 42,88 cm di lokasi lain di zona Kaffa. Selain itu, Fufa ( 2016)) mengamati genotipe jintan hitam dengan tinggi tanaman terpendek dan tertinggi masing-masing 41,83 dan 58,68 cm, dan 20 genotipe memiliki tinggi tanaman rata-rata keseluruhan 52,78 cm. Hal ini mencerminkan adanya variasi di antara genotipe yang diuji. Variasi dalam hasil biji dapat disebabkan oleh lingkungan, genetik, atau interaksi di antara kedua faktor ini (Rahimmalek et al., 2009 ; Shafie et al., 2009 ). Kehadiran variabilitas di antara genotipe jintan hitam disaksikan oleh berbagai macam sifat yang dipelajari. Hal ini menunjukkan bahwa ada kemungkinan untuk meningkatkan sifat-sifat ini melalui seleksi. Fufa ( 2016 ) melaporkan hasil biji sebesar 1,47, 0,92, dan 0,79 t ha −1 di Sinana, Ginir, dan Goro (semuanya di Ethiopia), masing-masing. Ermias et al. ( 2015 ) melaporkan rata-rata hasil biji sebesar 612 kg ha −1 untuk tiga varietas unggul jintan hitam yang dibudidayakan oleh petani di lokasi percobaan Kaya Kela, Pusat Penelitian Pertanian Bonga, Ethiopia. Hal ini menunjukkan bahwa plasma nutfah jintan hitam di Ethiopia memiliki rentang variasi yang luas untuk hasil biji. Hasil penelitian Emran et al. ( 2017 ) pada tiga puluh tujuh aksesi menunjukkan perbedaan yang nyata untuk semua karakter, termasuk hasil biji per tanaman, selama tahun 2014. Selama tahun 2015, aksesi-aksesi tersebut berbeda secara nyata untuk sebagian besar karakter kecuali tinggi tanaman dan biji per kapsul. Selama tahun 2014. Selama tahun 2015, aksesi-aksesi tersebut berbeda secara nyata untuk sebagian besar karakter kecuali tinggi tanaman dan biji per kapsul. Aksesi dengan hasil tertinggi memberikan 3,35 g per tanaman, sedangkan aksesi dengan hasil terendah memberikan 1,44 g selama tahun 2014. Selain itu, Asefa dan Beriso ( 2020 ) melaporkan bahwa rata-rata total hasil biji genotipe di seluruh lingkungan berkisar antara 2,45 hingga 1,61 t ha −1 . Rata-rata untuk jumlah kapsul per tanaman, biomassa, dan cabang primer berkisar antara 10,72 hingga 6,96, 63,74 hingga 39,79, dan 4,67 hingga 3,33, berturut-turut. Yewubdinber ( 2020 ) melakukan penelitian terhadap 25 genotipe jintan hitam di Pusat Penelitian Pertanian Debre-Zeit pada tahun 2018/2019. Hasil dari ANOVA mengungkapkan variasi genotipe untuk hasil biji berada di antara 1,86 dan 2,87 t ha −1 dengan rata-rata 2,29 kg ha −1 . Selain itu, Basazinew et al. ( 2023 ) melaporkan bahwa ANOVA gabungan di kedua lokasi mengungkapkan efek yang signifikan untuk genotipe dalam semua sifat kuantitatif kecuali jumlah cabang primer per tanaman. Variabilitas genotipe untuk hari hingga matang menciptakan peluang besar untuk mengembangkan varietas yang matang lebih awal dengan meningkatkan sifat-sifat yang terkait dengan hari hingga matang. Hasil ini akan membantu pemulia untuk mengembangkan varietas yang lebih baik dan sesuai untuk berbagai agroekologi di negara tersebut. Saeed et al. ( 2016)) di Iran menunjukkan perbedaan yang signifikan antara ekotipe untuk tinggi tanaman, jumlah cabang, jumlah kapsul, hari hingga awal berbunga, hari hingga berbunga penuh, berat 1000 biji, dan tingkat pengisian biji. Kisaran hari hingga berbunga dan matang akan membantu memilih galur yang matang awal untuk penggunaan di masa mendatang. Genotipe terendah membutuhkan waktu 23 hari untuk berbunga. Indeks panen tertinggi berkisar antara 8,54% hingga 35,85% (Iqbal et al., 2019 ). Variasi yang signifikan antara bahan jintan hitam yang diuji untuk sifat agro-morfologi yang diukur menunjukkan adanya tingkat variabilitas fenotipik yang tinggi, dan ini menyiratkan ketersediaan potensi besar untuk perbaikan jintan hitam di masa mendatang.
4.2 Estimasi fenotipik dan GCV
GCV dan PCV berkisar dari 3,81% hingga 49,84% dan 6,50% hingga 50,90%, masing-masing, untuk 16 sifat dari 64 genotipe jintan hitam. Salesh et al. ( 2010 ), Bharathiveeraman et al. ( 2012 ), Nwangburuka et al. ( 2012 ), dan Swati et al. ( 2014 ) menyimpulkan bahwa PCV dan GCV merupakan indikasi pengaruh yang lebih rendah dari faktor lingkungan dalam ekspresi sifat dan peluang yang lebih tinggi untuk meningkatkan sifat melalui pemuliaan. Demikian pula, Gholizade et al. ( 2019 ) melaporkan GCV dan PCV yang tinggi untuk jumlah kapsul dan PCV sedang untuk jumlah cabang primer. Preeti et al. ( 2019 ) juga melaporkan GCV dan PCV yang tinggi untuk jumlah kapsul. Tewodros et al. ( 2018 ) juga melaporkan estimasi PCV dan GCV yang rendah untuk hari hingga 50% pembungaan, hari hingga matang, tinggi tanaman, dan berat 1000 biji. Nilai GCV yang lebih rendah daripada nilai PCV untuk sifat-sifat tersebut mencerminkan variasi yang tampak tidak hanya karena efek genotipe tetapi juga keterlibatan efek lingkungan (Gholizade et al., 2019 ; Misra et al., 2013 ; Sarkar et al., 2015 ). Analisis variabilitas yang dilakukan untuk 10 karakter dalam 16 genotipe jintan hitam yang beragam mengungkapkan GCV dan PCV yang tinggi untuk cabang sekunder per tanaman, jumlah gabah per buah, jumlah buah per tanaman, panjang buah (cm), dan lebar buah (cm) (Singh & Singh, 2018 ). Selain itu, Yewubdinber ( 2020 ) melaporkan GCV dan PCV untuk 10 sifat yang diperkirakan dalam kisaran antara 3,91% dan 48,36% dan 6,90% dan 48,94%, masing-masing. Estimasi GCV dan PCV yang tinggi hingga sedang diamati untuk jumlah kapsul, jumlah cabang sekunder, dan jumlah kapsul. Basazinew et al. ( 2023 ) mengindikasikan bahwa tingkat variabilitas untuk sifat tertentu adalah wajib untuk program perbaikan tanaman apa pun, termasuk rempah-rempah. Hasil penelitian menunjukkan bahwa nilai estimasi GCV berkisar antara 0,66% hingga 18,41% untuk hari hingga matang dan hasil biji per plot, masing-masing. Demikian pula, nilai PCV berkisar antara 1,37% hingga 22,96% untuk hari hingga matang dan hasil biji per plot, masing-masing.
4.3 Estimasi heritabilitas dan GA
Dalam penelitian ini, heritabilitas arti luas ( H 2 ) yang diestimasi untuk 16 sifat dari 64 genotipe jintan hitam berkisar antara 19,18% (diameter kapsul) hingga 98,25% (untuk hasil biji per tanaman), sedangkan GA sebagai persentase rata-rata berkisar antara 4,67% (untuk diameter kapsul) hingga 100,70% (untuk indeks panen). Demikian pula, PhaniKrishna et al. ( 2015 ) menyarankan bahwa seleksi berdasarkan kinerja fenotipik genotipe akan efektif untuk meningkatkan sifat-sifat yang memiliki GA tinggi sebagai persentase rata-rata yang digabungkan dengan estimasi heritabilitas yang tinggi. Preeti et al. ( 2019 ) melaporkan heritabilitas sedang untuk hari hingga 50% pembungaan dan hari hingga matang. Selain itu, estimasi heritabilitas arti luas berkisar dari 30,17% untuk hasil biji per tanaman hingga 90,31% untuk hari hingga 50% pembungaan (Tewodros et al., 2018 ). Selain itu, penulis yang sama menunjukkan nilai heritabilitas tinggi untuk hari hingga 50% pembungaan (90,31%) dan hari hingga pembungaan pertama (87,34%). Estimasi sedang untuk durasi pembungaan (71,82%), berat 1000 biji (55,85), dan tinggi tanaman (47,48%). Di sisi lain, heritabilitas rendah dicatat untuk biomassa per plot (34,75%), hari hingga matang (31,75%), dan hasil biji per tanaman (30,17%) (Tewodros et al., 2018 ). ( 2018 ) menyelidiki hubungan timbal balik antara karakter kuantitatif dalam 13 aksesi biji hitam dalam kondisi tadah hujan di Pakistan dan melaporkan bahwa nilai heritabilitas lebih besar untuk cabang primer per tanaman, tinggi tanaman, jumlah kapsul per tanaman, jumlah biji per kapsul, berat 1000 biji, dan hasil biji. Demikian pula, Singh dan Singh ( 2018 ) melaporkan bahwa estimasi heritabilitas tinggi untuk jumlah biji per kapsul (0,86), jumlah kapsul per tanaman (0,78), panjang kapsul (0,64), hari hingga matang (0,59), jumlah cabang sekunder per tanaman (0,56), dan hasil per tanaman (0,47). Iqbal dkk. ( 2019 ) melaporkan bahwa estimasi heritabilitas dalam arti luas berkisar dari 0,28 untuk jumlah cabang hingga 0,98 untuk hari hingga matang. Heritabilitas tinggi (<50%) diamati untuk semua sifat kecuali jumlah cabang, yang menunjukkan ruang lingkup seleksi dari bahan ini. Induk yang beragam sifatnya dapat dipilih untuk memulai program hibridisasi dan pemuliaan. Preeti et al. ( 2019)) mencatat estimasi heritabilitas yang tinggi, dan estimasi GA yang tinggi sebagai persentase rata-rata diamati untuk hasil biji per tanaman, hasil biologis per tanaman, indeks panen, dan kapsul per tanaman, yang menunjukkan dominasi efek gen aditif untuk sifat-sifat ini. Estimasi heritabilitas tertinggi ditemukan untuk hasil biologis, hasil biji per tanaman, jumlah kapsul per tanaman, indeks panen, hari hingga berbunga, berat uji, dan tinggi tanaman. Karakter seperti hari hingga matang, jumlah cabang primer dan sekunder, dan biji per tanaman menunjukkan estimasi heritabilitas sedang. Ini menyiratkan bahwa seleksi dapat dilakukan untuk sifat-sifat ini karena mereka kurang dipengaruhi oleh lingkungan. GA tertinggi untuk biji per tanaman, diikuti oleh indeks panen, tinggi tanaman, dan hari hingga 50% berbunga. Keuntungan genetik tertinggi untuk hasil biji dan hasil biologis dan terendah untuk hari hingga matang dan hari hingga 50% berbunga. Yewubdinber ( 2020 ) menunjukkan bahwa heritabilitas arti luas ( H2 ) dan GA sebagai persentase rata-rata (GAM) berkisar antara 32,16 hingga 99,74 dan 4,58 hingga 98,61%, berturut-turut. Estimasi H2 dan GAM yang tinggi hingga sedang diamati untuk jumlah kapsul , jumlah cabang sekunder, dan jumlah kapsul. Basazinew dkk. ( 2023 ) menunjukkan bahwa heritabilitas arti luas untuk sifat-sifat kuantitatif yang dipelajari berkisar dari yang paling rendah 8,16% untuk jumlah biji per kapsul hingga yang tertinggi 89,47% untuk berat 1000 biji. Dengan demikian, nilai heritabilitas tinggi diperoleh pada berat 1000 biji (89,47%), tinggi tanaman (78,3%), hari hingga berbunga (76,65%), hari hingga mekar penuh (74,71%), jumlah kapsul per tanaman (66,86%), hasil biji per plot (64,33%), dan hari hingga 50% berbunga (62,56%). Hasil pendukung dilaporkan oleh Tewodros et al. ( 2018 ) pada hari jintan hitam hingga matang, hari hingga berbunga, dan hari hingga 50% berbunga. Heritabilitas sedang dalam arti luas tercatat pada jumlah biji per tanaman, hasil biji per hektar, jumlah cabang primer per tanaman, dan hasil biji per tanaman. Namun, nilai heritabilitas arti luas yang rendah ditunjukkan oleh hari hingga matang dan jumlah biji per kapsul. Tingginya H 2 akan menjadi korespondensi yang erat antara variasi genotipe dan fenotipe karena kontribusi lingkungan yang relatif kecil terhadap ekspresi fenotipe sifat tersebut (Singh et al., 1990 ).
4.4 Analisis komponen utama
Hasil serupa dilaporkan di Debre-Zeit bahwa empat komponen utama pertama mencakup sekitar 76,05% dalam 33 genotipe jintan hitam (Tewodros et al., 2018 ). Selain itu, Emran et al. ( 2017 ) melaporkan bahwa PCA hanya dapat mengekstraksi 84,18% dan 84,12% variabilitas di antara aksesi untuk semua sifat. Tiga PCA pertama dapat mengekstraksi 55,79% selama tahun 2014 dan 53,78 selama tahun 2015. Selama tahun 2014 dan 2015, PCA pertama mengekstraksi 21,41% dan 21,77%, PCA kedua mengekstraksi 18,66% dan 17,22%, dan PCA ketiga masing-masing 15,72% dan 14,79%. Ke-37 aksesi terpisah jauh selama kedua tahun tersebut, yang menunjukkan bahwa koleksi dari India dan Afghanistan jelas merupakan aksesi yang berbeda dengan lebih sedikit tumpang tindih genetik di antara mereka. Aksesi India dan Afghanistan menunjukkan pemisahan yang jelas dalam ruang PCA. Yewubdinber et al. ( 2022 ) melaporkan bahwa hasil PCA dari 12 sifat kuantitatif. PCA1 hingga PCA4 menyumbang persentase varians total yang bervariasi yang berkisar antara 9,98% hingga 28,43% dan menyumbang 70,62% dari total variasi. PCA1 menyumbang 28,43% terhadap total variabilitas di antara genotipe, sementara PCA2, PCA3, dan PCA4 masing-masing menunjukkan kontribusi 18,91%, 13,30%, dan 9,98% terhadap total variabilitas. Hari hingga pembungaan pertama (24,44%), hari hingga 50% pembungaan (23,50%), hari hingga 90% kematangan (21,64%), dan tinggi tanaman (9,02%) memiliki kontribusi lebih tinggi pada PCA1. Jumlah cabang sekunder (29,72%), diikuti oleh jumlah cabang primer, jumlah kapsul, dan jumlah biji per kapsul dengan masing-masing 23,54%, 21,52%, dan 10,99%, memiliki kontribusi lebih besar pada PCA2. Hasil biji (16,71%), jumlah biji per kapsul (13,34%), dan berat 1000 biji (39,33%) memiliki kontribusi yang relatif lebih besar pada varians total PCA4. Selain itu, Iqbal et al. ( 2013 ) menunjukkan bahwa sebagian besar variasi genetik dijelaskan oleh hari hingga kematangan, jumlah cabang per tanaman, jumlah kapsul per tanaman, biomassa, dan hasil biji pada PCA untuk sifat kuantitatif. Aksesi yang memiliki gen yang diinginkan untuk sifat-sifat yang penting secara ekonomi telah diidentifikasi dan karenanya direkomendasikan untuk digunakan secara langsung atau dimasukkan dalam program hibridisasi untuk pengembangan varietas. Selama tahun 2009, enam komponen utama terbentuk yang menyumbang 87,85% variabilitas, sementara selama tahun 2010, tiga PC terbentuk dan menyumbang 79,75% variabilitas. Untuk tahun 2011, lima PC terbentuk dan menyumbang 79,42% variabilitas. PC menunjukkan kontribusi sifat yang bervariasi di setiap tahun; hal ini mungkin disebabkan oleh kondisi iklim yang tidak seragam selama bertahun-tahun. Fluktuasi tinggi dari curah hujan musiman, suhu, laju penguapan, dan kelembaban memengaruhi hasil biji dan minyak pada tahun-tahun yang dilaporkan. Selain itu, Basazinew et al. ( 2023)) melaporkan bahwa empat komponen utama pertama menjelaskan 81,7% variasi yang ada di antara genotipe yang dipelajari. Hari hingga 50% berbunga, hari hingga matang, hari hingga mekar penuh, hari hingga berbunga, jumlah biji per tanaman, hasil biji per tanaman, dan jumlah kapsul per tanaman adalah sifat kontributor utama untuk variasi pada PC kedua, yang menyumbang 24,6% dari total variasi. Sifat kontributor utama untuk PC ketiga adalah variasi terutama dalam hasil biji per plot, hasil biji per hektar, jumlah kapsul per tanaman, jumlah biji per tanaman, dan hasil biji per tanaman. Selain itu, berat 1000 biji, tinggi tanaman, dan jumlah biji per kapsul adalah sifat kontributor utama untuk variasi pada PC keempat. Proporsi total varians fenotipik genotipe yang dijelaskan oleh PC ketiga dan keempat masing-masing sekitar 16,7% dan 8,5%. Oleh karena itu, PCA mengonfirmasi bahwa genotipe jintan hitam Ethiopia yang dikumpulkan memiliki keragaman tinggi, dan sebagian besar sifat yang dipertimbangkan tampaknya memiliki kontribusi tinggi terhadap variabilitas fenotipik total. Alboukadel ( 2017 ) menjelaskan bahwa sifat-sifat seperti hari hingga berbunga, hari hingga mekar penuh, hasil biji per tanaman, hari hingga matang, jumlah biji per tanaman, hari hingga 50% berbunga, dan tinggi tanaman merupakan kontributor utama terhadap variasi pada PC pertama, sedangkan hari hingga 50% berbunga, hari hingga matang, hari hingga mekar penuh, hari hingga berbunga, jumlah biji per tanaman, hasil biji per tanaman, dan jumlah kapsul per tanaman merupakan kontributor utama terhadap variasi pada PCA2. Sifat-sifat yang memiliki kontribusi lebih besar terhadap varians total setiap PCA menunjukkan bahwa sifat-sifat ini bertanggung jawab atas diferensiasi genotipe jintan hitam ke dalam kelompok yang berbeda.
4.5 Analisis Koefisien Korelasi
Korelasi karakter yang signifikan menunjukkan bahwa ada banyak ruang untuk seleksi langsung dan tidak langsung untuk perbaikan lebih lanjut. Korelasi signifikan diamati di antara hasil biji dan sifat lain yang dipelajari, tetapi korelasi positif yang sangat signifikan berkorelasi tinggi dan positif dengan hari pembentukan tunas, hari pembentukan kapsul, hari hingga 50% pembungaan, tinggi tanaman, dan kapsul per tanaman. Ini menunjukkan bahwa sifat-sifat ini adalah sifat penyumbang utama untuk perbaikan hasil biji pada genotipe jintan hitam. Oleh karena itu, karakter-karakter ini harus dipertimbangkan saat melakukan seleksi untuk perbaikan hasil pada nigella. Hasil serupa dilaporkan oleh Fufa ( 2016 ), Tewodros et al. ( 2018 ), Iqbal et al. ( 2019 ), Yewubdinber ( 2020 ), Zigyalew ( 2020 ), dan Basazinew et al. ( 2023)). Hari-hari pembentukan kuncup berkorelasi tinggi dan positif dengan hari-hari pembentukan kapsul, hari-hari hingga 50% pembungaan, hari-hari hingga kematangan, tinggi tanaman, cabang-cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar. Hari-hari hingga pembentukan kapsul berkorelasi tinggi dan positif dengan hari-hari hingga pembentukan kuncup, hari-hari hingga 50% pembungaan, hari-hari hingga kematangan, tinggi tanaman, cabang-cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar. Hari-hari hingga 50% pembungaan berkorelasi tinggi dengan hari-hari hingga pembentukan kuncup, hari-hari hingga pembentukan kapsul, hari-hari hingga kematangan, tinggi tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per hektar. Hari-hari hingga kematangan berkorelasi tinggi dengan hari-hari hingga pembentukan kuncup, hari-hari hingga pembentukan kapsul, hari-hari hingga 50% pembungaan, tinggi tanaman, cabang-cabang primer per tanaman, kapsul per tanaman, biji per kapsul, dan hasil biji per tanaman. Tinggi tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, cabang primer per tanaman, cabang sekunder per tanaman, kapsul per tanaman, panjang kapsul, biji per kapsul, dan hasil biji per hektar. Cabang primer per tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, tinggi tanaman, dan cabang sekunder per tanaman. Cabang sekunder per tanaman sangat berkorelasi dengan tinggi tanaman dan cabang primer per tanaman. Berat seribu biji sangat berkorelasi dengan hanya hasil biji per tanaman. Kapsul per tanaman sangat berkorelasi dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, tinggi tanaman, hasil biji per tanaman, dan hasil biji per hektar. Panjang kapsul sangat berkorelasi positif dengan tinggi tanaman dan diameter kapsul. Diameter kapsul sangat berkorelasi positif dengan panjang kapsul tetapi sangat berkorelasi negatif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, dan hari hingga kematangan. Biji per kapsul berkorelasi tinggi dan positif dengan hari pembentukan kuncup, hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, tinggi tanaman, dan hasil biomassa per hektar. Hasil biji per tanaman berkorelasi tinggi dan positif dengan hari hingga kematangan, berat 1000 biji, kapsul per tanaman, dan hasil biomassa per hektar. Hasil biomassa per hektar berkorelasi tinggi dan positif dengan biji per kapsul dan hasil biji per tanaman. Indeks panen berkorelasi tinggi dan negatif dengan hari pembentukan kapsul, hari hingga 50% pembungaan, hari hingga kematangan, dan hasil biomassa per hektar. Hasil serupa dilaporkan oleh Fufa ( 2018 ), Tewodros et al. ( 2018 ), Preeti et al. ( 2019 ), Iqbal et al. ( 2019 ), Yewubdinber ( 2020 ), Zigyalew ( 2020)), dan Basazinew et al. ( 2023 ).
4.6 Analisis data kualitatif
Demikian pula, percobaan direkam untuk percobaan yang dilakukan untuk menilai keragaman 24 genotipe jintan hitam dengan mempertimbangkan 10 parameter kualitatif yaitu elastisitas batang, warna bunga, pematangan biji, ukuran folikel, laju pembungaan, waktu pembungaan, luas daun, kerapatan cabang, daya tumbuh, dan ketebalan batang. Berdasarkan indeks keragaman Shannon–Weaver ( H ′), semua sifat dikategorikan tinggi kecuali warna bunga (Mehri et al., 2022 ). Dalam aspek ini, nilai H ′ untuk sebagian besar karakter fenotipik yang diamati menunjukkan tingkat keragaman normal di antara aksesi jintan hitam yang diuji, yang berkisar dari 0,47 untuk warna bunga hingga 1,58 untuk daya tumbuh, dengan nilai rata-rata 1,02 (Mehri et al., 2022 ). Hasilnya menunjukkan bahwa genotipe yang dikelompokkan dalam klaster yang sama memiliki kesamaan untuk banyak sifat tetapi perbedaan dengan genotipe lain dalam klaster lain dengan satu atau lebih sifat. Analisis klaster memisahkan genotipe ke dalam klaster, yang menunjukkan homogenitas tinggi dalam klaster dan heterogenitas tinggi antar klaster (Jaynes et al., 2003 ).
5 KESIMPULAN
Hasil penelitian ini menunjukkan adanya variasi yang signifikan untuk semua sifat yang dipelajari di antara genotipe jintan hitam yang dikumpulkan dari berbagai wilayah di Ethiopia. Performa rata-rata genotipe mengungkapkan rentang yang lebih lebar antara nilai minimum dan maksimum dari semua sifat kuantitatif kecuali panjang dan diameter kapsul; ini menunjukkan adanya variasi yang signifikan di antara genotipe jintan hitam yang termasuk dalam penelitian ini. Estimasi GCV, PCV, H 2 , dan GAM yang tinggi hingga sedang diamati untuk semua parameter kecuali hari pembentukan kuncup, hari hingga matang, panjang kapsul, dan diameter kapsul. Sekitar 76,05% variasi yang terjadi di antara genotipe yang dipelajari dijelaskan oleh lima komponen utama pertama. PCA mengkonfirmasi adanya variabilitas fenotipik yang tinggi di antara genotipe jintan hitam Ethiopia. Mengenai data kualitatif, aksesi yang dievaluasi menunjukkan keragaman yang tinggi kecuali untuk warna bunga, tipe pertumbuhan tanaman, dan rambut batang. Dalam analisis klaster untuk sifat kualitatif untuk 64 aksesi diklasifikasikan menjadi lima kelompok. Keragaman yang diamati di antara genotipe jintan hitam merupakan peluang untuk meningkatkan produktivitas tanaman baik melalui seleksi maupun hibridisasi untuk menggabungkan sifat-sifat yang diinginkan dari berbagai genotipe ke dalam keturunan segregasi atau hibrida. Selain itu, sejumlah sifat ekonomis utama jintan hitam, seperti hasil biji dan sifat-sifat yang terkait dengan hasil, dapat ditingkatkan melalui seleksi. Meskipun demikian, percobaan harus diulang di lebih banyak lokasi untuk mengonfirmasi temuan dan untuk mendapatkan informasi terkonsolidasi yang penting dalam program pemuliaan jintan hitam.



Tinggalkan Balasan