
ABSTRAK
Padang rumput perkotaan (UG) merupakan salah satu jenis infrastruktur hijau yang paling melimpah di kota-kota di seluruh dunia. Padang rumput ini menyediakan berbagai layanan ekosistem dan menjalankan banyak fungsi ekosistem. Untuk memastikan multifungsi UG, peningkatan keanekaragaman hayatinya sangat penting; namun, karena dampak urbanisasi dan pengelolaan yang tidak tepat, UG sering kali memiliki kekayaan spesies yang rendah. Meskipun vegetasi perkotaan telah dipelajari secara umum, wawasan ekologis terperinci tentang pola atribut fungsional, ekologis, dan komposisi vegetasi di berbagai ukuran, lokasi, dan jenis UG masih kurang. Pengetahuan ekologi yang lebih baik tentang vegetasi UG akan memfasilitasi pengelolaan yang bertujuan untuk meningkatkan keanekaragaman hayati dan multifungsinya. Di kota Wrocław, Eropa Tengah, kami mensurvei 244 petak UG yang berbeda dalam ukuran (kecil, sedang, dan besar), jenis (halaman rumput, tanggul, tepi jalan, dan UG di taman), dan lokasi (pusat kota dan pinggiran). Komposisi sifat fungsional, distribusi relung ekologi, dan ukuran keanekaragaman hayati dinilai berdasarkan komposisi spesies tanaman dan informasi yang diperoleh dari basis data terbuka, termasuk nilai indikator gangguan untuk tanaman. Hasil penelitian menunjukkan bahwa trade-off utama dalam komposisi sifat fungsional dan distribusi relung spesies terkait dengan hilangnya biomassa yang terkait dengan frekuensi pemotongan. Faktor-faktor yang diperiksa (ukuran petak, lokasi, dan jenis) secara signifikan memengaruhi vegetasi UG, dengan masing-masing memiliki pengaruh yang berbeda pada keanekaragaman hayati dan multifungsi. Banyak faktor yang terkait dengan perluasan perkotaan (misalnya, pulau panas perkotaan, perubahan reaksi tanah, fragmentasi lanskap, invasi biologis) sulit untuk dikurangi. Namun, pengelolaan yang tepat, seperti pemotongan yang luas, dapat membantu memoderasi dampak negatif urbanisasi. Meningkatkan atau mempertahankan proporsi forb yang tinggi dalam vegetasi juga akan meningkatkan multifungsi UG.
1 Pendahuluan
Di seluruh dunia, orang-orang semakin banyak tinggal di kota, yang mengarah pada proses urbanisasi global. Ruang yang diciptakan oleh pertumbuhan daerah perkotaan terutama melayani kebutuhan ekonomi warga, tetapi juga menyebabkan kerusakan ekosistem alami dan semi-alami (McKinney 2006 ). Sebuah kota mengembangkan lingkungan perkotaan yang berbeda dengan mengubah kondisi abiotik lokal, misalnya, suhu udara yang lebih tinggi dan polusi, kebisingan, kalsifikasi tanah, salinisasi, dan gangguan (McDonnell dan Pickett 1990 ). Dari perspektif lanskap, dominasi pembangunan atas infrastruktur hijau menyebabkan isolasi spasial habitat, yang pada gilirannya membatasi penyebaran benih tanaman dan perekrutan bibit (untuk tinjauan lihat de Barros Ruas et al. 2022 ). Rangkaian kondisi ini memungkinkan pertimbangan urbanisasi sebagai gradien ekologis (McDonnell dan Pickett 1990 ).
Gradien urbanisasi menentukan banyak faktor fisik dan kimia yang membentuk komposisi sifat fungsional dan distribusi relung ekologi tanaman tergantung pada kekuatan faktor-faktor utama (Williams et al. 2015 ; Aronson et al. 2016 ). Di satu sisi, urbanisasi mengarah pada homogenisasi kumpulan tanaman dengan menyaring spesies tanaman yang tidak dapat bertahan dalam kondisi perkotaan; di sisi lain, lanskap perkotaan yang dinamis menyebabkan pergantian yang tinggi dalam komunitas tanaman dalam skala yang relatif kecil (Kühn dan Klotz 2006 ; Knapp et al. 2008 ; Lokatis dan Jeschke 2022 ). Daerah perkotaan menampung spesies tanaman yang berasal dari habitat yang berbeda secara ekologis dan geografis yang datang secara spontan atau tidak sengaja, atau secara sengaja diperkenalkan (Kühn dan Klotz 2006 ; McKinney 2006 ; Lososová et al. 2012 ). Akibatnya, keanekaragaman spesies tanaman di kota meningkat, menciptakan campuran spesies baru dan khusus perkotaan yang tidak ditemukan di luar wilayah dengan tekanan manusia yang kuat (Kowarik 2011 ). Untuk mengembangkan praktik pengelolaan yang memastikan masa depan kota yang berkelanjutan dan beragam hayati, memahami peran relatif filter abiotik, biotik, dan antropogenik yang membentuk komunitas tanaman lokal adalah penting (Aronson et al. 2016 ).
Di antara tipe habitat terbuka yang paling umum di kota-kota di seluruh dunia adalah padang rumput perkotaan (UG) (Ignatieva et al. 2020 ). Istilah ini awalnya merujuk pada halaman rumput klasik yang sering dipotong (untuk ulasan, lihat Ignatieva et al. 2020 ), tetapi baru-baru ini berkembang dan sekarang mencakup semua tipe vegetasi terbuka yang lebih atau kurang sering dipangkas di area perkotaan (Klaus 2013 ), bahkan vegetasi non-rumput yang didominasi forb (Smith et al. 2015 ). Di UG, pemotongan terjadi dengan intensitas yang bervariasi tergantung pada fungsi yang dilayani, lokasi tertentu, dan ketersediaan dana pengelolaan. Area yang paling sering dipangkas termasuk halaman rumput kota, taman bermain, dan tepi jalan, yang dipotong beberapa kali dalam setahun (Ignatieva et al. 2015 ). Area yang kurang dipangkas secara intensif termasuk sisa-sisa padang rumput semi-alami, fragmen alang-alang dan vegetasi alang-alang tinggi di dekat badan air dan sungai, dan lahan kosong dengan berbagai kondisi habitat. Pengelolaan yang ekstensif mengganggu suksesi sekunder dan sangat penting untuk mempertahankan UG dari semua jenis terlepas dari kondisi lingkungan suatu lokasi (Klaus dan Kiehl 2021 ).
UG, sebagai bagian dari infrastruktur hijau kota, harus multifungsi dan dengan demikian melayani jangkauan seluas mungkin dalam penyediaan, pengaturan, dan layanan budaya, serta menciptakan habitat perlindungan bagi spesies padang rumput (Ignatieva et al. 2020 ; Klaus dan Kiehl 2021 ). Hubungan antara keanekaragaman hayati dan multifungsi ekosistem tidaklah langsung (Meyer et al. 2018 ), tetapi dalam kasus padang rumput, peningkatan kekayaan spesies meningkatkan multifungsi mereka (Schils et al. 2022 ), terutama dalam menghadapi perubahan iklim dan intensifikasi penggunaan lahan (Allart et al. 2024 ). Selain dampak kekayaan spesies, komposisi spesies tumbuhan dan sifat-sifatnya memengaruhi layanan ekosistem yang disediakan. Selain itu, komunitas dengan keanekaragaman fungsional yang lebih tinggi, yang sering berkorelasi dengan kekayaan spesies, lebih tahan terhadap invasi biotik (Frankow-Lindberg 2012 ).
Menilai ekologi UGs melalui pendekatan yang berfokus pada ciri fungsional tanaman memungkinkan mengungkap dan menggeneralisasi mekanisme yang membentuk keanekaragaman hayati dan respons komunitas tanaman terhadap faktor lingkungan (Lavorel dan Garnier 2002 ). Pengetahuan tentang ciri fungsional tanaman mana yang akan diuntungkan oleh urbanisasi, dan mana yang tidak, sangat penting untuk memperkirakan layanan ekosistem dan melestarikan keanekaragaman hayati di dunia yang semakin terurbanisasi (Williams et al. 2015 ).
Perbedaan vegetasi antara UG dapat disebabkan oleh kondisi lingkungan setempat, serta jenis dan/atau frekuensi gangguan. Vegetasi juga dapat bervariasi karena pengaruh lanskap sekitarnya, yang diakibatkan oleh lokasi dan ukuran petak UG. Tujuan utama dari penelitian ini adalah untuk menguji hipotesis berikut: (1) UG yang berbeda dalam jenis (pinggir jalan, tanggul sungai, halaman rumput, dan UG di taman kota), lokasi (pusat kota dan pinggiran), dan ukuran akan berbeda dalam komposisi sifat fungsional tanaman, distribusi relung ekologi spesies, dan keanekaragaman hayati. (2) Distribusi sifat dan relung ekologi berkorelasi dengan pola keanekaragaman taksonomi dan fungsional. Untuk menguji hipotesis ini, kami mengumpulkan data tentang komposisi spesies tanaman dari 244 petak UG yang tersebar di kota berukuran sedang di Eropa Tengah. Selain itu, pengetahuan ekologi yang diperoleh dari hasil kami tentang pola karakteristik vegetasi UG juga dirujuk ke rekomendasi pengelolaan yang ditujukan untuk meningkatkan multifungsi UG.
2 Bahan dan Metode
2.1 Wilayah Studi
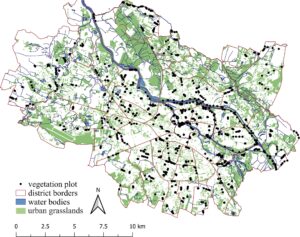
Penelitian ini dilakukan di kota Wrocław yang terletak di wilayah Silesia, Polandia, Eropa Tengah, pada ketinggian berkisar antara 105 hingga 156 mdpl. Wrocław meliputi area seluas sekitar 300 km 2 dan berpenduduk sekitar 900.000 jiwa. Suhu tahunan rata-rata adalah 9,7°C, bulan terhangat adalah Juli (19,9°C), dan terdingin adalah Januari (−0,5°C). Curah hujan tahunan rata-rata adalah 548 mm.
Ruang hijau perkotaan mencakup 53% dari wilayah kota, dan UG mencakup ca. 9523 ha (32% dari wilayah kota). Petak padang rumput kecil dengan ukuran hingga 0,5 ha adalah yang paling banyak (Mollashahi et al. 2020 ). UG mewakili berbagai area hijau yang dikelola secara formal dan informal dengan intensitas pengelolaan yang bervariasi, termasuk halaman rumput hias, halaman rumput rekreasi, area rekreasi, area layanan, padang rumput di perumahan multi-keluarga, tepi jalan, padang rumput di tanggul sungai, dan area kosong. Intensitas pemotongan tergantung pada keputusan preferensi pemilik atau pengelola lahan. Jika diterapkan, campuran benih yang paling umum digunakan termasuk Festuca rubra , Lolium perenne , dan Poa pratensis (informasi pribadi otoritas kota, pengamatan sendiri). Padang rumput semi-alami di pinggiran dan lahan kosong hanya dipangkas sekali setiap beberapa tahun atau bahkan puluhan tahun, sedangkan halaman rumput hias dapat dipangkas hingga beberapa minggu sekali (Dewan Penghijauan Kota Wrocław, komunikasi pribadi, pengamatan sendiri).
2.2 Desain Penelitian dan Pengumpulan Data
Skema pengambilan sampel dirancang berdasarkan peta UG yang diperoleh dari Basis Data Objek Topografi Polandia (BDOT10k) ( https://www.geoportal.gov.pl/pl/dane/baza-danych-obiektow-topograficznych-bdot10k ). Hanya padang rumput dengan akses tak terbatas, termasuk padang rumput publik, semi-publik, dan semi-privat (misalnya, yang berada di perumahan multi-keluarga) yang diperiksa. UG dikategorikan menurut ukuran: kecil (S; luas < 0,5 ha); sedang (M; luas 0,5–10 ha); dan besar (L; luas > 10 ha). Berdasarkan lokasi dan jumlah warga yang terdaftar, kami membedakan UG di pusat kota (C) dan pinggiran kota (P) berdasarkan Mollashahi et al. ( 2020 ). Terakhir, kami mengenali empat jenis UG dengan fungsi yang berbeda: pinggir jalan (R), tanggul sungai (E), padang rumput di taman (Pa), dan halaman rumput (L). Sebagai halaman rumput, kami mempertimbangkan halaman rumput hias, area layanan, area kosong, dan padang rumput di perumahan multi-keluarga, tidak termasuk yang terkait dengan jalan, sungai, atau taman. Setiap petak ditentukan berdasarkan ukuran, lokasi, dan jenisnya; untuk ringkasan lihat Tabel S1 .
Empat titik pengambilan sampel yang didistribusikan secara merata ditetapkan di setiap petak padang rumput, dan selama kerja lapangan di setiap titik pengambilan sampel, plot vegetasi (1 × 1 m) dibuat untuk mencatat spesies tanaman vaskular dan persentase tutupannya. Kerja lapangan dilakukan selama musim tanam (dari Mei hingga September) pada tahun 2020 dan 2021. Plot vegetasi yang tutupan lapisan pohon atau semaknya > 30% dan/atau tutupan lapisan herba < 50% dikeluarkan dari analisis. Selain itu, semua spesies pohon dan semak dikeluarkan dari basis data. Akhirnya, 938 plot dari 244 petak padang rumput dianalisis (Gambar 1 ). Nomenklatur tanaman disatukan menurut daftar Euro+Med 2006+ ( 2021 ).
Untuk mengidentifikasi distribusi relung ekologi spesies, nilai indikator Ellenberg (IV) rata-rata dihitung (Ellenberg et al. 1992 ) menggunakan basis data Pladias (Chytrý et al. 2021 ). Dengan demikian, nilai indikator untuk frekuensi pemotongan rumput (nilai indikator gangguan) (Midolo et al. 2022 ) dihitung.
Untuk setiap petak UG, rata-rata tutupan spesies yang tercatat dihitung. Ciri fungsional dan nilai indikator untuk setiap petak UG dihitung dengan rata-rata nilai (rata-rata tertimbang komunitas [CWM]; Garnier et al. 2004 ), yang ditimbang berdasarkan kelimpahan relatif spesies.
Kami menghitung jumlah semua spesies (kekayaan spesies, SR) dan ukuran keanekaragaman Rao dari ciri-ciri spesies fungsional. Dengan demikian, jumlah spesies asli (asli dan arkeofit), spesies asing (neofit termasuk spesies invasif dan hias), spesies invasif, dan jumlah spesialis padang rumput dihitung. Daftar spesies tanaman dengan kategori yang ditetapkan disajikan dalam Tabel S3 .
Matriks korelasi peringkat r s Spearman antara variabel yang digunakan dalam analisis ditunjukkan pada Tabel S4 .
2.3 Analisis Statistik
Untuk memperoleh gambaran umum komposisi spesies dari vegetasi yang diteliti, skala multidimensi non-metrik (NMDS) diterapkan, berdasarkan jarak Bray–Curtis. Sebelum NMDS, data tentang kelimpahan spesies ditransformasikan dengan Hellinger, kemudian ditransformasikan dengan Wisconsin ganda.
Untuk menyederhanakan struktur data, kami menerapkan analisis komponen utama (PCA) secara terpisah untuk komposisi fungsional tanaman dan untuk diferensiasi relung ekologi. Jumlah sumbu PCA yang dianalisis ditetapkan menggunakan pendekatan ‘tongkat patah’. Untuk analisis lebih lanjut, kami menggunakan pemuatan PCA dari kelompok variabel sebagai variabel komposit. Sebelum PCA, data distandarisasi dengan mengurangi rata-rata dan membaginya dengan deviasi standar untuk setiap nilai setiap variabel. Dengan demikian, untuk mencapai gambaran yang lebih baik dari hasil yang diperoleh, kami membatasi spesies tanaman yang paling mewakili gradien PCA. Prosedur sederhana berikut diterapkan: 20 UG dengan nilai skor tertinggi dari PCA tertentu dan 20 UG dengan nilai skor terendah dari PCA tertentu ditetapkan. Kemudian, spesies dengan frekuensi tertinggi dalam kelompok UG ditentukan.
Untuk mengeksplorasi perbedaan antara UG dari berbagai kategori dalam komposisi fungsional tanaman, diferensiasi relung ekologi yang tercermin dari pemuatan PCA, dan ukuran keanekaragaman hayati, kami menerapkan kerangka kerja model linear umum (GLM). Keluarga model Gaussian untuk nilai Rao, sifat dan skor PCA IV, quasi-Poisson untuk kekayaan spesies, quasi-binomial untuk proporsi spesies asli, dan binomial negatif untuk proporsi kelompok spesies padang rumput, asing, dan invasif digunakan. Untuk distribusi Gaussian, kami menerapkan fungsi tautan ‘identitas’; untuk quasi-Poisson dan binomial negatif, fungsi ‘log’; dan untuk quasibinomial, ‘logit’. Kami menggunakan model glm dasar dari paket ‘stats’, dan model nol yang digelembungkan dari paket ‘pscl’ (Tim Inti R 2023 ; Jackman 2024 ) untuk data dengan jumlah nilai nol yang berlebihan (fraksi spesies asing, invasif, dan padang rumput). Diagnostik model dilakukan menggunakan paket ‘kinerja’ (Lüdecke et al. 2021 ) dengan memeriksa kelebihan atau kekurangan dispersi data dan homogenitas serta distribusi residual, serta dengan inspeksi visual plot residual. Kami menguji efek utama pengelompokan variabel serta interaksi di antara variabel tersebut, menerapkan fungsi ANOVA dalam paket ‘mobil’ dengan uji ‘F’ (Fox dan Weisberg 2019 ). ANOVA tipe III diterapkan saat interaksi terdeteksi, dan ANOVA tipe II saat hanya ditemukan efek utama. Uji post hoc dilakukan dengan uji Tukey dalam paket ‘multicomp’ dan ‘emmeans’, dengan koreksi Bonferroni (Lenth 2018 ).
Korelasi peringkat Spearman rs dihitung di antara nilai skor UG yang diperoleh dari PCA untuk komposisi fungsional tanaman, dan diferensiasi relung ekologi, serta ukuran keanekaragaman hayati (Tabel 2 ). Semua perhitungan dilakukan dalam R versi 4.2.2 (Tim Inti R 2023 ).
3 Hasil
3.1 Struktur Vegetasi
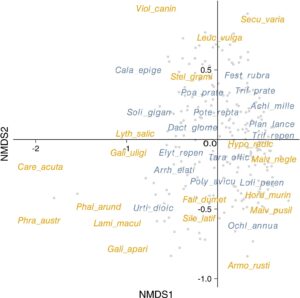
Sebanyak 327 spesies tumbuhan vaskular telah tercatat. Di antara spesies-spesies ini, 254 adalah spesies asli (termasuk archaeophytes), yang 30 spesiesnya secara ekologis merupakan karakteristik padang rumput semi-alami. Sebanyak 36 spesies asing, termasuk 21 spesies invasif, telah tercatat (Tabel S3 ). Ada 68 spesies graminoid dan 259 spesies forb. Di antara taksa yang paling sering adalah Lolium perenne (hadir pada 62,4% dari petak UG), Taraxacum officinale agg. (57,1%), dan Elytrigia repens (56,7%) (Tabel S3 ). Komposisi keseluruhan spesies dalam UG disajikan pada grafik NMDS (Gambar 2 ). Jumlah semua spesies per UG berkisar dari 3 hingga 37 spesies (rata-rata = 17,8, SD = 7,03), dan SR berkorelasi positif dengan nilai Rao (Tabel S4 ).
3.2.1 Pengurangan Dimensi PCA untuk Komposisi Fungsional
Pertukaran utama dalam variabilitas komposisi sifat spesies dapat digambarkan sebagai gradien pesaing versus ruderal (Ciri PC 1, 32,7% dari varians yang dijelaskan), forb versus graminoid (Ciri PC 2, 12,0%), gradien stres (Ciri PC 3, 10,8%), dan fraksi terofit dalam vegetasi (Ciri PC 4 9,8%).
Sumbu pertama (Sifat PC 1) menggarisbawahi kontras antara vegetasi yang didominasi oleh spesies kompetitif dengan nilai tinggi kandungan bahan kering daun dan tinggi, versus vegetasi yang didominasi oleh ruderal tahunan (Tabel 1 ). Spesies tanaman khas dalam kelompok pertama adalah spesies dengan toleransi rendah terhadap pemotongan ( Calamagrostis epigejos , Elytrigia repens , Urtica dioica , Solidago gigantea , dan Phalaroides arundinacea ); sedangkan, untuk kelompok kedua adalah Ochlopoa annua , Lolium perenne , dan Hordeum murinum , semuanya beradaptasi dengan gangguan tanah. Sifat PC 1 berkorelasi negatif dengan nilai Rao dan proporsi spesies asli; itu berkorelasi positif dengan proporsi spesies padang rumput (Tabel 2 ).
| Sifat PC 1 | Sifat PC 2 | Sifat PC 3 | Sifat PC 4 | Bahasa Indonesia: IV PC 1 | Bahasa Indonesia: Komputer IV 2 | Bahasa Indonesia: Komputer IV 3 |
|---|---|---|---|---|---|---|
| Pesaing versus orang-orang kasar | Forbs versus graminoid | Menekankan | Terofit versus hemikriptofit | Frekuensi pemotongan | Reaksi tanah | Suhu |
| LDMC (0,33) | Forb (0,5) | Toleransi terhadap stres (0,54) | Terofit (0.46) | Memotong rumput (0.55) | Reaksi tanah (0.71) | Suhu (0.89) |
| Tinggi tanaman (0,31) | Autokori (0.23) | Autokori (0.44) | Massa benih (0,32) | Cahaya (0,51) | Salinitas (0,48) | Ringan (0.23) |
| Abadi (0.31) | Hemerokori (0.22) | Bunga kamofit (0,27) | Graminoid (0,22) | Salinitas (0,22) | Ketersediaan N (0,34) | Reaksi tanah (0.22) |
| Geofit (0.31) | Perkiraan tanggal kedaluwarsa (0.21) | Hidrokori (0,23) | Hidrokori (0,22) | Kelembaban tanah (−0,42) | Kelembaban tanah (0,28) | Salinitas (−0,2) |
| Kompetitif (0.31) | Dua tahunan (0.21) | Hidrofit (0.22) | Hidrofit (0.22) | Ketersediaan N (−0,45) | Memotong rumput (0.21) | Kelembaban tanah (−0,21) |
| Klonalitas (0.24) | Anemokori (−0,22) | Anemokori (−0,25) | Anemokori (−0,21) | |||
| Hidrofit (0,2) | Hemikriptofit (−0,24) | Zookori (−0,25) | Forb (−0,22) | |||
| Hemikriptofit (−0,23) | Hidrokori (−0,28) | Kompetitif (−0,25) | Chamaephyta (−0,29) | |||
| Tahunan (−0,31) | Graminoid (−0,5) | Hemikriptofit (−0,32) | ||||
| Ruderal (−0,33) | ||||||
| ve = 32,7 | ve = 12,0 | ve = 10,8 | ve = 9,8 | sudah = 39,1 | sudah = 24,2 | saya = 15,3 |
Catatan: Warna latar belakang menunjukkan kekuatan korelasi positif (palet hangat) dan negatif (palet dingin). Untuk penyederhanaan, hanya ciri dengan nilai terbesar (> 0,2) atau nilai terendah (< 0,2) yang ditampilkan. Untuk semua variabel, lihat Tabel S5 . Singkatan: LDMC, kandungan bahan kering daun; SLA, luas daun spesifik; ve, varians yang dijelaskan (%).
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sifat PC 1 | 1 | ||||||||||||
| Sifat PC 2 | 2 | ||||||||||||
| Sifat PC 3 | 3 | ||||||||||||
| Sifat PC 4 | 4 | -0,15 | |||||||||||
| Bahasa Indonesia: IV PC 1 | 5 | -0,77 | -0,40 | -0,26 | |||||||||
| Bahasa Indonesia: Komputer IV 2 | 6 | -0,30 | -0,23 | 0.19 | 0.20 | ||||||||
| Bahasa Indonesia: Komputer IV 3 | 7 | -0,24 | 0.21 | -0,53 | 0.29 | ||||||||
| Kekayaan | 8 | 0.21 | 0.22 | -0,20 | -0,48 | ||||||||
| Rao | 9 | -0,19 | 0.62 | -0,27 | 0.27 | 0.42 | |||||||
| Proporsi spesies padang rumput | 10 | 0.23 | 0.12 | 0.29 | -0,15 | -0,18 | -0,29 | -0,14 | 0.32 | ||||
| Proporsi spesies asli | 11 | 0.39 | -0,37 | 0,15 | -0,45 | ||||||||
| Proporsi spesies asing | 12 | -0,39 | 0.36 | -0,15 | 0.46 | -0,99 | |||||||
| Proporsi spesies invasif | 13 | -0,45 | 0.35 | -0,16 | 0.44 | -0,16 | -0,90 | 0,90 |
Catatan: Untuk penyederhanaan, hanya korelasi signifikan yang ditampilkan. Warna latar belakang menunjukkan kekuatan korelasi positif (palet hangat) dan negatif (palet dingin).
Sumbu kedua (Sifat PC 2) menggambarkan gradien dominasi forb dalam vegetasi dengan forb ( Urtica dioica , Galium aparine , Solidago gigantea , Taraxacum officinale agg., tetapi juga Elytrigia repens ) versus graminoid ( Lolium perenne , Festuca rubra , Poa pratensis , Elytrigia repens , tetapi juga Taraxacum officinale agg. dan Trifolium repens ) (Tabel 1 ). Menariknya, Elytrigia repens dan Taraxacum officinale agg. paling sering muncul di UG dengan nilai tertinggi dan terendah dari skor Sifat PC 2. Sifat PC 2 berkorelasi positif dengan semua kekayaan spesies, nilai Rao, dan proporsi spesies padang rumput (Tabel 2 ).
Sumbu ketiga (Sifat PC 3) merepresentasikan gradien dominasi spesies autokorik yang toleran terhadap stres. Sumbu berkorelasi positif dengan cakupan tinggi oleh spesies padang rumput khas yang toleran terhadap pemotongan (seperti Arrhenatherum elatius , Dactylis glomerata , Galium mollugo ) atau genangan ( Phragmites australis , Urtica dioica ). UG dengan nilai terendah dari skor Sifat PC 3 dicirikan oleh frekuensi tinggi Solidago gigantea dan Calamagrostis epigejos (spesies dengan resistensi rendah terhadap pemotongan dan genangan). Selain itu, Elytrigia repens , Urtica dioica , Poa pratensis , dan Festuca rubra sering muncul di UG dengan nilai tertinggi dan terendah dari skor Sifat PC 3 (Tabel 1 ). Sifat PC 3 berkorelasi positif dengan semua kekayaan spesies dan proporsi padang rumput dan spesies asli; Padahal, hal ini berkorelasi negatif dengan proporsi spesies asing dan spesies invasif (Tabel 2 ).
Sumbu keempat (Ciri PC 4) menggambarkan kontras antara vegetasi dengan tutupan terofit yang tinggi dengan massa biji yang tinggi (misalnya, Ochlopoa annua , Hordeum murinum ) versus hemicryptophytes ( Plantago lanceolata , Trifolium repens , Achillea millefolium , Lolium perenne ) (Tabel 1 ). Dalam UG, spesies dengan skor yang ditempatkan pada kedua sisi Ciri PC 4 sering muncul, termasuk Lolium perenne , Taraxacum officinale agg., dan Dactylis glomerata . Ciri PC 4 berkorelasi positif dengan proporsi spesies asing dan proporsi spesies invasif; sedangkan itu berkorelasi negatif dengan jumlah semua spesies dan proporsi padang rumput dan spesies asli (Tabel 2 ).
3.2.2 Reduksi Dimensi PCA untuk Distribusi Relung Ekologi
Variabilitas utama dalam distribusi relung ekologi yang diperkirakan dengan bioindikasi (nilai indikator Ellenberg [IV]) dapat diartikan sebagai hasil dari gradien frekuensi pemotongan (IV PC1, 39,1% dari varians yang dijelaskan), reaksi tanah, dan salinitas (IV PC2, 24,2%), dan suhu (IV PC3, 15,3%) (Tabel 1 ).
Sumbu pertama (IV PC 1) menggambarkan gradien antara vegetasi UGs yang didominasi oleh tanaman yang membutuhkan cahaya dan tahan terhadap frekuensi pemotongan yang tinggi ( Lolium perenne , Taraxacum officinale agg., Trifolium repens ) versus vegetasi UGs dengan tutupan tanaman yang membutuhkan nitrogen dan kelembaban yang tinggi ( Urtica dioica , Galium aparine , Phalaroides arundinacea ). IV PC 1 berkorelasi positif dengan proporsi spesies asli; sedangkan berkorelasi negatif dengan proporsi spesies padang rumput, invasif, dan asing (Tabel 2 ).
Sumbu kedua (IV PC 2) menggambarkan gradien reaksi tanah yang tinggi, salinitas, dan ketersediaan nutrisi. Urtica dioica , Phalaroides arundinacea , Phragmites australis , Dactylis glomerata , dan Lolium perenne paling sering muncul di UG dengan nilai IV PC 2 tertinggi; sedangkan, Festuca rubra , Agrostis capillaris , Calamagrostis epigejos , Elytrigia repens , dan Poa pratensis paling sering muncul di UG dengan nilai IV PC 2 terendah. Sumbu ini berkorelasi negatif dengan semua kekayaan spesies, nilai Rao, dan proporsi spesies padang rumput (Tabel 2 ).
Sumbu ketiga (IV PC 3) menggambarkan gradien suhu. UG dengan skor tertinggi untuk sumbu ketiga paling sering ditempati oleh Lolium perenne , Elytrigia repens , Ochlopoa annua , Solidago gigantea , Urtica dioica , dan Hordeum murinum . UG dengan nilai skor terendah dari IV PC3 didominasi oleh Festuca rubra , Elytrigia repens , Poa pratensis , Achillea millefolium , dan Dactylis glomerata . IV PC 3 berkorelasi positif dengan nilai Rao dan proporsi spesies asing dan invasif; sedangkan, berkorelasi negatif dengan proporsi padang rumput dan spesies asli (Tabel 2 ).
Semua nilai pemuatan variabel untuk sumbu komponen utama yang dibedakan, dan persentase variasi yang dijelaskan oleh sumbu disajikan dalam Tabel S5 .
3.3 Hasil GLM
Hasil dari semua model GLM ditunjukkan pada Tabel S6 , sedangkan statistik deskriptif untuk kelompok ada pada Tabel S7 . Dalam teks, demi kesederhanaan, hanya hasil yang signifikan secara statistik yang disajikan.
3.3.1 Hasil GLM untuk Komposisi Fungsional Tanaman
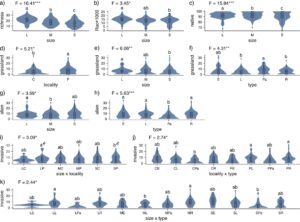
Uji GLM (Tabel S6 ) menunjukkan perbedaan dalam nilai Trait PC 1 (pesaing vs. ruderal) di antara UG dengan tipe yang berbeda: tanggul dicirikan oleh nilai yang lebih tinggi dibandingkan dengan tepi jalan dan halaman rumput (Gambar 3a ). Kami mengamati interaksi lokalitas dan ukuran dalam kasus Trait PC 1. Biasanya, petak-petak yang terletak di pinggiran memiliki nilai yang lebih tinggi dibandingkan dengan petak-petak yang ditempatkan di pusat kota, dengan pengecualian petak-petak besar yang memiliki nilai yang sama terlepas dari lokasinya (Gambar 3b ). UG dengan fungsi yang berbeda berbeda secara signifikan sehubungan dengan nilai Trait PC 3 mereka: taman memiliki nilai yang lebih tinggi dibandingkan dengan halaman rumput dan tepi jalan (Gambar 3c ).

Hasil GLM (Tabel S6 ) mengungkapkan perbedaan signifikan antara UG dari kategori lokasi yang berbeda dalam nilai skor IV PC 1 (frekuensi pemotongan). UG di tengah dicirikan oleh nilai IV PC 1 yang tinggi; sedangkan, UG di pinggiran dicirikan oleh nilai IV PC 1 yang rendah (Gambar 4a ). Ada juga perbedaan sehubungan dengan jenis UG: halaman rumput dan tepi jalan memiliki nilai yang lebih tinggi daripada padang rumput di tanggul sungai dan di taman (Gambar 4b ). Kami juga menemukan perbedaan dalam nilai skor IV PC 2 (reaksi tanah) untuk UG dari lokasi yang berbeda: UG di tengah dicirikan oleh nilai IV PC 2 yang lebih tinggi, dan UG di pinggiran, dengan nilai yang lebih rendah (Gambar 4c ).

3.3.3 Hasil GLM untuk Pengukuran Keanekaragaman Hayati
Ukuran petak UG memiliki efek signifikan pada kekayaan spesies total: jumlah spesies yang diinangi meningkat seiring dengan ukuran petak (Gambar 5a ). Kami mengamati pola yang sama sehubungan dengan proporsi spesies asli (meningkat dengan ukuran petak) (Gambar 5c ). Juga, indeks keanekaragaman fungsional (Rao) lebih tinggi pada petak besar dibandingkan dengan petak UG kecil (Gambar 5b ). Proporsi spesies padang rumput dipengaruhi oleh lokalitas, ukuran, dan jenis UG, masing-masing (tanpa interaksi antar kelompok): spesialis padang rumput menciptakan fraksi jumlah spesies yang lebih tinggi di pinggiran dibandingkan dengan pusat kota (Gambar 5d ), serta di petak UG besar dibandingkan dengan yang kecil (Gambar 5e ). Selain itu, kami menemukan bahwa UG di taman menjadi tuan rumah bagi fraksi spesialis padang rumput yang lebih tinggi dibandingkan dengan halaman rumput dan tepi jalan (Gambar 5f ). Kekayaan spesies asing lebih tinggi pada petak besar dibandingkan pada petak berukuran sedang (Gambar 5g ), sementara itu lebih rendah di UG di taman dibandingkan dengan semua tipe yang tersisa (Gambar 5h ). Dalam kasus spesies invasif, kami mendeteksi efek signifikan yang terkait dengan interaksi: ukuran × lokasi, lokasi × tipe, dan ukuran × tipe. Hasil pengujian post hoc untuk efek interaksi ukuran × lokasi tidak mendeteksi perbedaan signifikan pada p < 0,05, tetapi ada kontras terkuat antara bercak UG kecil dan besar yang ditempatkan di pinggiran ( nilai p untuk uji post hoc: p = 0,077): di pinggiran, vegetasi bercak besar mengandung lebih banyak spesies invasif daripada bercak kecil (Gambar 5i ); namun, ukuran efeknya kecil ( d Cohen = 0,152). Penyerbu lebih sering terjadi di halaman rumput dan tepi jalan yang ditempatkan di pinggiran dibandingkan dengan UG di taman di pusat kota (Gambar 5j ). Halaman rumput besar dan tepi jalan sedang menampung spesies invasif yang lebih proporsional dibandingkan dengan halaman rumput sedang dan UG kecil di taman (Gambar 5k ).
Kami mencatat jumlah spesies tanaman yang relatif tinggi, yaitu 327 spesies di UG di Wrocław. Jumlah spesies yang relatif tinggi kemungkinan besar disebabkan oleh lanskap perkotaan yang dinamis yang menyebabkan pergantian kumpulan spesies dalam skala spasial yang kecil, sehingga memungkinkan keberadaan spesies ruderal, segetal, dan riparian, termasuk spesies non-asli (Tabel S3 ).
Hasil kami mendukung hipotesis bahwa komposisi fungsional tanaman, diferensiasi relung ekologi, dan ukuran keanekaragaman hayati berbeda di seluruh ukuran petak UG, lokasi, dan jenis. Efek dari faktor-faktor tertentu pada vegetasi selaras dengan pengetahuan umum tentang ekologi perkotaan. Karakteristik vegetasi mencerminkan rezim pengelolaan khusus yang terkait dengan jenis UG (Ignatieva dan Hedblom 2018 ), ukuran petak, dan gradien urbanisasi dari pusat ke pinggiran, yang semuanya memengaruhi struktur vegetasi, termasuk kekayaan spesies (McDonnell dan Pickett 1990 ; Aronson et al. 2016 ; Horak et al. 2022 ). Kami juga dapat mendukung hipotesis kedua bahwa indeks keanekaragaman hayati tanaman vaskular di UG berkorelasi dengan pola distribusi relung ekologi dan komposisi sifat fungsional. Namun, korelasi ini, bersama dengan efek interaksi di antara faktor-faktor yang dipelajari pada vegetasi, jauh lebih jarang dieksplorasi (misalnya, Williams et al. 2015 ).
4.1 Interpretasi Ekologis terhadap Gradien yang Diamati dan Hubungannya dengan Indeks Keanekaragaman Hayati
Hasilnya menunjukkan bahwa gangguan (termasuk frekuensi pemotongan) adalah faktor utama yang mempengaruhi trade-off dalam komposisi sifat fungsional (Sifat PC 1) dan diferensiasi relung ekologi (IV PC 1). Vegetasi UG di pusat memiliki kelimpahan tanaman yang lebih besar yang beradaptasi dengan pemotongan yang sering daripada UG di pinggiran (Gambar 4a ), karena gradien urbanisasi umum (biasanya menurun dari pusat ke pinggiran). Dengan demikian, halaman rumput dan tepi jalan dipangkas lebih sering daripada UG di taman dan di tanggul sungai (Gambar 4b ), yang muncul dari fungsi mereka yang berbeda. Habitat paling produktif di sepanjang sungai jarang dipangkas, karena mereka mewakili vegetasi riparian semi-alami yang dikelola untuk pengendalian erosi dan mitigasi kerusakan banjir (Prus et al. 2018 ). Ini juga dapat membantu menjelaskan korelasi negatif antara nilai indikator frekuensi pemotongan dan IV untuk nitrogen (Tabel S4 ).
Indikator frekuensi pemotongan sangat berkorelasi dengan indikator gangguan tanah (lihat Bagian 2.3 ), yang menunjukkan bahwa gradien IV PC 1 dapat mencerminkan tingkat gangguan keseluruhan yang memengaruhi habitat. Biasanya, penurunan frekuensi pemotongan bermanfaat untuk kekayaan spesies di UG karena meratakan gangguan (Rudolph et al. 2017 ; Chollet et al. 2018 ; Sehrt et al. 2020 ; Watson et al. 2020 ). Peningkatan frekuensi pemotongan (IV PC1) berkorelasi positif dengan proporsi spesies asli dan berkorelasi negatif dengan proporsi padang rumput dan spesies invasif. Kami berasumsi bahwa pemotongan mengurangi efek kompetitif spesies invasif yang kehadirannya biasanya mengurangi kekayaan spesies (Pyšek et al. 2010 ).
Gradien gangguan juga dapat dikaitkan dengan PC 1 dalam kasus sifat spesies. Vegetasi yang dipangkas secara ekstensif dan tidak terganggu didominasi oleh spesies abadi yang tinggi dan kompetitif, dengan siklus hidup yang tidak terkait dengan hilangnya biomassa yang menghasilkan biomassa tinggi, karena biomassa biasanya berkorelasi dengan tinggi tanaman (Axmanová et al. 2012 ). Kelompok sifat ini terkait dengan penurunan keragaman fungsional (Tabel 2 ). Dominasi semusim dan ruderal pada vegetasi yang diamati di sayap berlawanan dari gradien Sifat PC 1 terutama terlihat di halaman rumput, serta petak kecil dan sedang di pusat kota (Gambar 3a,b ), yang dipengaruhi oleh tingkat gangguan yang tinggi. Hasil ini konsisten dengan temuan Deák et al. ( 2016 ), yang melaporkan bahwa habitat yang lebih terpapar urbanisasi menampung spesies yang lebih toleran terhadap gangguan.
Perbedaan dalam fraksi ruderal dan/atau tanaman tahunan di berbagai jenis UG (Ciri PC 1) dapat dikaitkan dengan invasi biologis. Invasi tanaman lebih mungkin terjadi di lingkungan yang terganggu (misalnya, gangguan tanah) dan pada tahap awal suksesi (Catford et al. 2012 ; Richardson et al. 2000 ). Namun, kecenderungan sebaliknya diamati di sini: fraksi tanaman tahunan dan ruderal yang menunjukkan gangguan tanah berkorelasi positif dengan proporsi spesies asli (Tabel 2 ). Kami mengaitkan hal ini dengan potensi penyebaran spesies asing yang masih terbatas di lanskap perkotaan: ruderal asli lebih sering daripada yang bukan asli, mungkin karena kehadirannya di bank benih tanah.
Dominasi tumbuhan perdu (Ciri PC 2) berkorelasi positif dengan kekayaan spesies dan keanekaragaman fungsi (Tabel 2 ). Dalam kondisi perubahan iklim, UG dengan proporsi tumbuhan perdu yang sama dengan rumput akan memberikan layanan ekosistem yang lebih kuat (Rojas-Botero et al. 2023 ). Dari sudut pandang praktis, hasil penelitian menunjukkan bahwa peningkatan tutupan tumbuhan perdu menguntungkan multifungsi UG.
Reaksi langsung vegetasi terhadap polusi tanah yang terkait dengan tingkat urbanisasi, biasanya menurun dari pusat ke pinggiran, diwakili oleh gradien kedua dalam distribusi relung ekologi (IV PC 2). Tanah perkotaan lebih basa dan mengandung kadar garam lebih tinggi daripada tanah di daerah sekitarnya (misalnya, Kida dan Kawahigashi 2015 ; Mónok et al. 2021 ). Kandungan kalsium tanah yang lebih tinggi di pusat kota sebelumnya ditunjukkan untuk Wrocław (Mollashahi et al. 2023 ). Akibatnya, vegetasi UG di pusat menunjukkan nilai gradien IV PC 2 yang lebih tinggi, terlepas dari jenis dan ukurannya. Biasanya, penggunaan umum garam pencair es memengaruhi vegetasi pinggir jalan dengan mempromosikan halofit (Fekete et al. 2022 ), tetapi kurangnya reaksi signifikan padang rumput di tepi jalan dalam kasus kami dapat dikaitkan dengan pembatasan penggunaan garam dalam beberapa tahun terakhir, ditingkatkan oleh musim dingin yang hangat. Umumnya, tanah perkotaan dianggap miskin nitrogen dibandingkan dengan tanah non-perkotaan (Guilland et al. 2018 ). Hasil kami tidak mengonfirmasi ekspektasi ini; bahkan sebaliknya, spesies dengan nilai IV yang lebih tinggi untuk nitrogen lebih melimpah di pusat kota (Gambar 4c ). Gradien urbanisasi tanah berkorelasi negatif dengan kekayaan spesies, keanekaragaman fungsional, dan proporsi spesies padang rumput (Tabel 2 ).
Plot dengan nilai Trait PC 4 yang tinggi (pertukaran antara vegetasi pada tahap awal suksesi dan vegetasi yang mengalami tekanan injakan) dapat diharapkan untuk mengkarakterisasi UG kecil atau yang terletak di pusat kota karena benih yang berat biasanya dianggap sebagai adaptasi untuk hidup di lingkungan perkotaan yang berbahaya (Williams et al. 2015 ). Kami tidak mengamati perbedaan seperti itu, dan itu dapat disebabkan oleh sifat-sifat vegetasi yang dipelajari: kami berfokus pada UG dan mengabaikan area dengan tanah yang baru saja terganggu (misalnya, tanah terlantar, tempat bangunan).
Penyortiran spesies tanaman relung sepanjang gradien suhu (IV PC 3) terkait dengan dua aspek urbanisasi: pulau panas perkotaan dan pengenalan spesies asing—yang sebagian besar berasal dari rentang asli yang lebih hangat atau lebih kering (Géron et al. 2021 ). Hasilnya tidak mengonfirmasi bahwa UG dari pusat kota berbeda dari yang ada di pinggiran dalam nilai sepanjang gradien suhu. Namun, kami mengamati frekuensi spesies yang membutuhkan hangat yang lebih tinggi di UG di tanggul sungai daripada di UG di taman, terutama di petak yang lebih kecil (Gambar 4d ).
Trait PC 3 (pertukaran antara vegetasi yang didominasi oleh pesaing dan vegetasi yang didominasi oleh penoleransi stres) tampaknya secara langsung dihasilkan dari toleransi terhadap pemotongan rumput dan genangan air; namun, tidak seperti Trait PC 1, hal itu tidak melibatkan jenis gangguan lain (misalnya, gangguan tanah). Dalam kasus kami, spesies yang toleran terhadap stres terdiri dari spesies rumput yang khas untuk padang rumput semi-alami yang digunakan secara ekstensif dan spesies riparian non-padang rumput yang toleran terhadap genangan air. Para pesaing mewakili tanaman asli yang invasif dan ekspansif yang mampu menghentikan proses suksesi tetapi tidak toleran terhadap genangan air dan pemotongan rumput. UG dengan nilai tinggi mengenai sumbu ini memiliki fraksi spesies asli dan spesialis padang rumput yang tinggi, sementara mereka rendah dalam spesies asing dan invasif. Hasilnya menggarisbawahi peran padang rumput di taman (Gambar 3c ) yang dipangkas secara ekstensif tetapi teratur dan relatif aman dari gangguan tanah yang khas dari halaman rumput dan tepi jalan. Selain itu, vegetasi UG di taman dicirikan oleh jumlah spesies asing yang rendah (Gambar 5h ). Akibatnya, vegetasinya menyusun kembali sebagian besar vegetasi padang rumput semi-alami (Gambar 5f ). Hal ini juga dapat dikaitkan dengan sejarah petak-petak tertentu: taman-taman tersebut biasanya dibuat di pinggiran kota bersejarah dengan vegetasi pedesaan dan kesinambungan habitat yang relatif panjang.
Hasil kami mengonfirmasi bahwa pengaruh ukuran petak merupakan salah satu faktor terpenting yang memengaruhi kekayaan spesies (Heegaard et al. 2007 ). Dalam ‘pemandangan kota’, ukuran besar area hijau sering dikaitkan dengan peningkatan kekayaan spesies tanaman (Angold et al. 2006 ; Matthies et al. 2017 ; Horak et al. 2022 ). Selain kekayaan spesies, vegetasi petak yang lebih besar juga memiliki keragaman fungsional yang lebih besar, dan terdiri dari proporsi spesies asli yang lebih tinggi, termasuk spesialis padang rumput. Karena korelasi positif antara kekayaan spesies, keragaman fungsional, dan berbagai layanan ekosistem yang diberikan (Cadotte et al. 2011 ; Schils et al. 2022 ; Allart et al. 2024 ; Rojas-Botero et al. 2023 ), studi kami menyoroti peran petak UG besar dalam konservasi keanekaragaman hayati asli dan pemberian layanan ekosistem. Selain itu, vegetasi yang lebih kaya spesies dengan redundansi fungsional yang lebih besar tampaknya lebih stabil dan tangguh terhadap gangguan dalam layanan ekosistem yang disediakan (Biggs et al. 2020 ). Pola fraksi spesies invasif dalam vegetasi menarik: perhatian yang rendah diberikan pada pemeliharaan petak-petak besar di pinggiran mengakibatkan jumlah penyerbu yang lebih tinggi dibandingkan dengan petak-petak kecil. Hasilnya menunjukkan bahwa invasi biologis dapat menjadi ancaman bagi pemeliharaan keanekaragaman hayati UG di kota-kota dan harus dikendalikan. Sering dinyatakan bahwa vegetasi perkotaan dapat dianggap sebagai ekosistem baru, kadang-kadang bahkan disebut ‘sifat tipe keempat’ yang terdiri dari campuran spesies asli dan asing. Namun, harus digarisbawahi bahwa tidak semua spesies asing bersifat invasif, dan pengendalian penyerbu, karena dampak lingkungannya, harus dipertahankan di kota-kota (untuk tinjauan, lihat Teixeira dan Fernandes 2020 ).
4.2 Keterbatasan Penelitian
Bahasa Indonesia: Dalam studi kami, kami mengambil sampel dari sejumlah besar petak UG (untuk tinjauan data yang tersedia lihat Fekete et al. 2024 ) yang dimiliki oleh agen yang berbeda. Akibatnya, memperoleh informasi terperinci tentang rezim pemotongan dan riwayat gangguan untuk semua petak yang diperiksa terbukti tidak layak. Sebaliknya, kami menggunakan bioindikasi, yang menawarkan keuntungan yang jelas tetapi juga menyajikan batasan tertentu, seperti interkorelasi antara nilai indikator tertentu (Diekmann 2003 ). Secara khusus, kami mengalami masalah dengan nilai indikator dari frekuensi pemotongan dan gangguan tanah, yang sangat berkorelasi dalam kasus vegetasi yang diperiksa. Meskipun demikian, kami berasumsi bahwa kami dapat menangkap efek dari pendorong terpenting dari struktur vegetasi UG, dan hasil yang disajikan di sini juga berlaku untuk kota-kota lain yang terletak di lingkungan yang sama, setidaknya di Eropa. Pada akhirnya, kami fokus pada multifungsi UG yang dihasilkan dari kekayaan spesies. UG menyediakan layanan yang berbeda, yang dapat saling eksklusif (misalnya, nilai olahraga vs. pemeliharaan keanekaragaman hayati), dan beberapa layanan dapat dianggap sebagai layanan yang merugikan, tergantung pada konteksnya (Von Döhren dan Haase 2015 ). Dengan demikian, fungsi utama dari suatu kawasan tertentu juga harus dipertimbangkan.
4.3 Implikasi dan Kesimpulan Manajemen
Elemen-elemen tertentu dari ekosistem kota secara negatif memengaruhi multifungsi UG dan akan sangat sulit untuk dinegasikan, termasuk efek pulau panas perkotaan, pengenalan spesies non-asli, polusi tanah, dan fragmentasi lanskap. Meskipun demikian, hasilnya menunjukkan bahwa komposisi fungsional tanaman UG sebagian besar dibentuk oleh gangguan, termasuk frekuensi pemotongan (Ciri PC 1) dan oleh proporsi tumbuhan perdu yang terbatas (Ciri PC 2). Fraksi tumbuhan perdu yang rendah tidak terkait dengan kumpulan spesies yang rendah di kota (259 spesies ditemukan) tetapi lebih pada kemungkinan terbatas untuk migrasi dan pendirian (Piana et al. 2019 ; Mollashahi et al. 2020 ). Hasil yang diperoleh di sini menunjukkan bahwa peningkatan fraksi tumbuhan perdu dapat dilakukan dalam ukuran, lokasi, dan jenis UG apa pun karena mereka tidak berbeda sehubungan dengan gradien ini. Dengan demikian, praktik pengelolaan tertentu dapat menguntungkan kekayaan spesies dan keanekaragaman fungsional, dengan demikian meningkatkan multifungsi vegetasi UG dan ketahanannya terhadap faktor-faktor yang terkait dengan perluasan kota. Praktik ini meliputi peningkatan luas UG dengan tetap menjaga petak yang ada, penyemaian ulang tanaman herba, dan penyesuaian waktu pemotongan agar bertepatan dengan produksi benih tanaman.
Struktur vegetasi UG, yang dapat atau harus diubah (misalnya, melalui peningkatan atau pengurangan frekuensi pemotongan, atau penambahan campuran benih yang sesuai), dapat dengan mudah diidentifikasi oleh ahli botani terlatih, tetapi tidak harus oleh pekerja di perusahaan pengelolaan tanaman hijau atau perencana kota. Dengan demikian, penting bagi pemerintah kota untuk membuka posisi bagi ahli biologi perkotaan, yang akan mampu menyiapkan rencana pengelolaan yang memberikan manfaat bagi masyarakat dan alam setempat. Menurut pendapat kami, mustahil untuk menerapkan solusi berbasis alam tanpa pengetahuan profesional tentang organisme hidup dan proses di antara mereka.



Tinggalkan Balasan