
Abstrak
- Menghitung hewan saat populasi dikumpulkan secara spasial (misalnya, koloni pengembangbiakan atau bersarang, lokasi persinggahan atau tempat berkumpul) meningkatkan akurasi dan efisiensi upaya survei untuk estimasi kelimpahan. Ortomosaik yang dihasilkan dari citra drone umumnya digunakan untuk menghitung populasi yang dikumpulkan, tetapi penghitungan ini rentan terhadap kesalahan deteksi yang sering kali diabaikan dalam estimasi kelimpahan.
- Termotivasi oleh kebutuhan akan protokol pemantauan untuk peristiwa bersarang massal Penyu Sungai Amerika Selatan Raksasa ( Podocnemis expansa ), kami mengembangkan pendekatan pemodelan baru untuk memperkirakan kelimpahan populasi satwa liar yang terkumpul secara spasial menggunakan hitungan berbasis drone dalam ortomosaik sambil memperhitungkan berbagai sumber kesalahan. Kami menggunakan kombinasi data penandaan-penglihatan ulang dan jumlah populasi keseluruhan untuk memperhitungkan: (i) populasi terbuka selama peristiwa bersarang; (ii) individu yang tidak dapat dideteksi selama penerbangan; (iii) hitungan ganda karena proses pembangunan ortomosaik; (iv) individu yang ditandai terdeteksi dalam mosaik dengan tanda yang tidak dapat diidentifikasi.
- Dari data penandaan-penemuan, kami memperkirakan bahwa probabilitas bersarang harian adalah 0,37, dan bahwa 35% individu yang menggunakan gundukan pasir pada malam hari hadir selama penerbangan drone pagi hari. Kami juga menemukan bahwa 20% kura-kura yang berjalan di orthomosaic adalah hitungan ganda, dan bahwa probabilitas mengidentifikasi tanda di karapas adalah 0,78. Total ukuran populasi diperkirakan sekitar ~41.000 kura-kura selama 12 hari musim bersarang, menandai agregasi kura-kura air tawar terbesar di dunia saat ini. Dengan membandingkan pendekatan kami dengan estimasi kelimpahan berdasarkan model yang lebih sederhana dan dengan hitungan tanah visual, kami menunjukkan manfaat pendekatan kami dan pentingnya memperhitungkan berbagai sumber kesalahan saat menghitung hewan di orthomosaic.
- Sintesis dan aplikasi . Pendekatan yang dikembangkan dapat diterapkan pada beberapa konteks untuk mensurvei populasi yang terkumpul secara spasial secara efisien menggunakan orthomosaik yang berasal dari drone, dan untuk memahami fenologi di lokasi agregasi ini. Kami memberikan rekomendasi umum untuk merencanakan survei dan membahas implementasi pendekatan kami menggunakan jenis metode penandaan dan asumsi model lainnya.
1. PENDAHULUAN
Kelimpahan adalah variabel fundamental dalam ekologi dan konservasi, misalnya, untuk mempelajari dinamika populasi, predator-mangsa dan interaksi interspesifik, serta untuk menilai dampak konversi habitat dan perubahan iklim global. Selain itu, dengan memantau kelimpahan dari waktu ke waktu, adalah mungkin untuk mendeteksi dan memprediksi tren populasi spesies buruan, invasif, atau terancam, bersama dengan menilai efektivitas tindakan pengelolaan untuk mengendalikan atau meningkatkan populasi ini (Butchart et al., 2010 ; Moussy et al., 2022 ). Namun, memperkirakan kelimpahan dapat menimbulkan tantangan yang signifikan, terutama di daerah yang luas dan ekstensif di mana spesies terjadi pada kepadatan rendah, sehingga sulit untuk mendeteksi individu dan mendapatkan jumlah yang akurat. Secara kebetulan, beberapa spesies satwa liar menunjukkan perilaku musiman di mana individu berkonsentrasi di daerah kecil untuk beristirahat, berinteraksi secara sosial, kawin, berkembang biak, dan/atau bersarang, memberikan peluang besar untuk menghitungnya (Brown, 2016 ). Misalnya, burung air berkumpul dalam koloni bersarang (Jovani et al., 2016 ; Rolland et al., 1998 ), anjing laut berkumpul di lokasi tempat mencari makan dan berkembang biak (Hoekendijk et al., 2023 ; Procksch et al., 2020 ), burung bersama-sama menggunakan lokasi persinggahan selama migrasi jarak jauh (Cohen et al., 2021 ; Schmaljohann et al., 2022 ), dan kura-kura secara bersamaan bersarang di gundukan pasir dan pantai (Forero-Medina et al., 2021 ; Scheelings, 2023 ). Oleh karena itu, menghitung hewan selama periode agregasi spasial ini dapat secara signifikan meningkatkan akurasi dan efisiensi upaya survei untuk memperkirakan dan memantau kelimpahan.
Baru-baru ini, survei berbasis drone telah muncul sebagai metode yang efisien dan kurang invasif untuk mengambil sampel populasi satwa liar yang dikumpulkan secara spasial (Christie et al., 2016 ; Linchant et al., 2015 ; Lyons et al., 2019 ). Menggunakan drone (juga dikenal sebagai kendaraan udara tak berawak, UAV; atau pesawat yang dipiloti dari jarak jauh, RPA) untuk menghitung individu yang dikumpulkan dari atas telah terbukti lebih akurat dan tepat dibandingkan dengan survei berbasis darat (Goebel et al., 2015 ; Hodgson et al., 2016 , 2018 ; Ratcliffe et al., 2015 ), sementara juga menyebabkan lebih sedikit gangguan pada hewan (Krause et al., 2021 ). Protokol umum yang digunakan untuk survei drone adalah merencanakan penerbangan dengan tumpang tindih tinggi antara foto-foto berurutan dan strip lateral, menggabungkan gambar-gambar yang dikumpulkan menjadi mosaik orthorektifikasi tunggal (yaitu orthomosaic; Westoby et al., 2012 ; Wolf et al., 2014 ). Saat mengambil sampel populasi agregat, penerbangan ini biasanya mencakup seluruh area tempat individu berkumpul (misalnya area koloni burung, Weinstein et al., 2022 ; atau pulau tempat anjing laut berkumpul, Procksch et al., 2020 ).
Namun, menghitung individu satwa liar dalam orthomosaik selama peristiwa agregasi ini tunduk pada beberapa sumber kesalahan yang tidak diinginkan, yang berpotensi membiaskan estimasi kelimpahan jika tidak ditangani dengan benar (Brack et al., 2018 ; Brack et al., 2025 ). Misalnya, seekor individu mungkin tidak dapat diamati dalam citra yang dikumpulkan (yaitu tidak tersedia untuk dideteksi) dengan tersembunyi di bawah vegetasi, di bawah air, atau sementara di luar area yang diterbangi (misalnya mencari makan di tempat lain). Selain itu, bahkan jika individu tersebut dapat diamati dalam gambar, pengamat manusia atau algoritma deteksi dapat gagal mendeteksinya. Lebih jauh lagi, hewan yang bergerak selama penerbangan drone dapat muncul beberapa kali di lokasi yang berbeda dalam foto yang digunakan untuk membuat orthomosaik (Gambar 1 ) (Brack et al., 2025 ). Akhirnya, karakteristik penting adalah bahwa agregasi ini umumnya bersifat sementara, dengan individu datang dan pergi selama beberapa hari, yang menyebabkan fluktuasi dalam ukuran populasi. Misalnya, selama musim bersarang, berkembang biak, atau migrasi burung dan anjing laut, individu dapat tiba dan meninggalkan area koloni pada hari yang berbeda sepanjang musim. Karakteristik “populasi terbuka” ini dapat menyebabkan estimasi kelimpahan yang bias jika tidak diperhitungkan. Yang mengkhawatirkan, kesalahan ini banyak diabaikan dalam estimasi kelimpahan yang diperoleh dari hitungan ortomosaik survei berbasis drone.

Dalam studi ini, kami bertujuan untuk mengembangkan pendekatan baru untuk memperkirakan kelimpahan populasi satwa liar yang teragregasi secara spasial menggunakan hitungan berbasis drone dalam orthomosaik sambil memperhitungkan berbagai sumber kesalahan. Pendekatan ini bergantung pada kombinasi dua jenis kumpulan data: penampakan individu yang ditandai dan jumlah populasi keseluruhan. Studi ini dimotivasi oleh kebutuhan untuk memperkirakan kelimpahan dan menguraikan protokol pemantauan untuk Kura-kura Sungai Raksasa Amerika Selatan ( Podocnemis expansa , selanjutnya disebut hanya sebagai “kura-kura sungai”) selama agregasi kura-kura air tawar terbesar yang diketahui di dunia. Setiap tahun selama musim kemarau, ribuan kura-kura sungai berkumpul untuk bersarang di gundukan pasir dan pantai Sungai Guaporé/Iténez, di sepanjang perbatasan Brasil-Bolivia (cekungan Amazon) (Gambar 2 ). Bahasa Indonesia: Sementara peristiwa bersarang massal ini dulunya umum di seluruh lembah Amazon dan Orinoco, populasi telah menurun drastis terutama karena eksploitasi berlebihan untuk konsumsi daging dan telur, dan agregasi besar ini sekarang langka (Forero-Medina et al., 2021 ). Metode sebelumnya untuk memperkirakan kelimpahan kura-kura sungai bergantung pada penghitungan tukik setelah mereka muncul (dan kemudian mengekstrapolasi jumlah betina menggunakan jumlah rata-rata telur per sarang) atau hitungan visual kura-kura dewasa dari tanah (Forero-Medina et al., 2021 ). Menghitung tukik adalah metode invasif dan memakan waktu karena memerlukan pagar perimeter gundukan pasir dan memanipulasi sejumlah besar tukik. Hitungan kura-kura dewasa secara visual di tanah juga menghadirkan keterbatasan karena kesulitan yang ditimbulkan oleh penghitungan ribuan individu yang saling menghalangi dan terus bergerak. Baru-baru ini, drone telah diuji untuk mensurvei populasi penyu sungai (Fagundes & Ferrara, 2022 ), yang sangat menjanjikan sebagai metode yang terstandarisasi, tepat, dan efisien untuk memperkirakan ukuran populasi selama peristiwa bersarang ini. Hal ini penting untuk menilai tren populasi dan efektivitas tindakan konservasi untuk spesies tersebut.

Kami menerapkan pendekatan pemodelan yang dikembangkan untuk memperkirakan populasi penyu sungai selama acara bersarang massal. Kami memperhitungkan berbagai sumber kesalahan, seperti ketersediaan individu, individu yang bergabung dan meninggalkan populasi lokal selama periode pengambilan sampel, dan penghitungan ganda karena proses pembuatan ortomosaik. Meskipun awalnya terinspirasi untuk meningkatkan pemantauan penyu sungai, kerangka kerja yang dikembangkan sangat serbaguna dan dapat dengan mudah digunakan atau diadaptasi ke beberapa konteks berbeda di mana populasi agregat disurvei menggunakan ortomosaik drone. Oleh karena itu, kami membahas aplikasi dan perluasan metode yang dikembangkan untuk skenario survei satwa liar lainnya.
2 BAHAN DAN METODE
2.1 Wilayah Studi
Penelitian ini dilakukan di Sungai Guaporé/Iténez, salah satu anak sungai utama Sungai Madeira, di cekungan Amazon (Gambar 2 ). Sungai Guaporé/Iténez membentang sekitar 1210 km, yang sebagian besar membentang di sepanjang perbatasan Brasil-Bolivia. Curah hujan tahunan di daerah tersebut berkisar antara 1500 dan 1600 mm, yang terbagi dalam dua musim yang berbeda, yaitu musim hujan (Desember–Mei) dan musim kemarau (Juni–November) (Pouilly et al., 2012 ). Selama musim air surut, gundukan pasir dan pantai yang besar muncul, yang digunakan oleh penyu sungai untuk bersarang.
Kura-kura Sungai Amerika Selatan Raksasa adalah salah satu kura-kura air tawar yang paling sosial (Ferrara et al., 2014 ), menyusuri sungai dalam kelompok besar dan berkumpul di depan gundukan pasir bersarang sekitar bulan Juli atau Agustus di wilayah Amazon ini. Kura-kura sungai betina bersarang secara serempak di gundukan pasir tertentu yang mereka pilih untuk tujuan ini (Alho & Pádua, 1982 ; Ferrara et al., 2010 ), dimulai saat permukaan air paling rendah (September hingga November). Peristiwa bersarang massal di Guaporé/Iténez adalah yang terbesar yang diketahui untuk spesies ini di seluruh wilayahnya (Forero-Medina et al., 2021 ). Sementara kura-kura sungai dapat menggunakan beberapa gundukan pasir untuk bersarang setiap tahun, kami memilih satu gundukan pasir tertentu pada tahun 2021 ( Praia da Ilha ) untuk mensurvei populasi (Gambar 2 ). Gumuk pasir ini adalah lokasi bersarang utama untuk tahun itu, memusatkan sebagian besar individu dan menyajikan bersarang massal terbesar. Penyu sungai meninggalkan air dan memasuki gundukan pasir biasanya pada malam hari (Vogt, 2008 ). Beberapa individu bersarang pada malam yang sama, sementara yang lain menjelajahi area tersebut untuk kembali pada malam yang berbeda untuk bersarang (Ferrara et al., 2023 ). Seekor individu dapat memasuki gundukan pasir beberapa kali sebelum bersarang, dan setelah bersarang, ia tidak kembali ke gundukan pasir (Ferrara et al., 2023 ).
2.2 Pengumpulan data
Survei drone dilakukan setiap hari antara 26 September dan 04 Oktober 2021, dimulai segera setelah matahari terbit, sekitar pukul 6 pagi. Kami menggunakan drone multirotor DJI Mavic 2 Enterprise Advanced yang membawa sensor tampak 48 Mpx. Untuk menutupi seluruh gundukan pasir, kami melakukan empat misi penerbangan berturut-turut yang memakan waktu total sekitar 1 jam per hari untuk menyelesaikannya. Kami memprogram penerbangan drone pada 50 m di atas permukaan tanah, dengan 80% tumpang tindih frontal dan 70% tumpang tindih lateral. Pengaturan penerbangan ini menghasilkan jarak pengambilan sampel tanah (GSD) 1 cm. Empat penerbangan setiap hari menghasilkan sekitar 1500 foto per hari. Sebelum setiap survei drone (sekitar pukul 3 pagi), kami menandai sekitar 100 individu yang berada di gundukan pasir, mengecat simbol unik di atas karapas mereka dengan cat putih, dengan tujuan untuk mengidentifikasi mereka nanti pada gambar drone (Gambar 2e ). Prosedur pengambilan sampel dilakukan berdasarkan nomor lisensi SISBIO/ICMBio 80087, yang mencakup otorisasi untuk kegiatan kerja lapangan di Sungai Guaporé dan penangkapan serta penandaan penyu.
Foto-foto yang dikumpulkan setiap hari dijahit menjadi orthomosaic harian menggunakan perangkat lunak OpenDroneMap™ ( https://www.ondronemap.org/ ) (Gambar 2c ). Dua pengamat meninjau setiap orthomosaic harian dalam perangkat lunak QGIS, menggunakan kisi untuk memandu pencarian dan membuat anotasi semua kura-kura sungai yang terdeteksi. Ketika kura-kura memiliki karapas yang ditandai, itu diidentifikasi jika memungkinkan atau diberi anotasi sebagai tanda yang tidak teridentifikasi (biasanya karena individu tersebut memiliki pasir di karapasnya). Selain itu, setiap deteksi (untuk individu yang tidak ditandai dan ditandai) diklasifikasikan menjadi bersarang atau berjalan (lihat Bagian 2.3 ). Individu yang bersarang dapat dibedakan dari yang berjalan karena mereka berada di area inti gundukan pasir, di dalam lubang di pasir dan dengan tubuh mereka dimiringkan ke bawah di ujung belakang. Kami tidak menyertakan data penampakan tanda untuk tanggal 30 September (yaitu kami hanya menggunakan jumlah keseluruhan) karena buruknya kualitas orthomosaic yang dihasilkan (cuaca yang sangat berawan menghasilkan foto yang gelap) menghalangi identifikasi tanda.
Kami mengorganisasikan data penandaan-penemuan kembali ke dalam tiga subset, satu matriks penangkapan-penangkapan kembali multistatus konvensional dan dua jenis hitungan harian individu yang ditandai ini. Subset pertama terdiri dari riwayat perjumpaan setiap individu yang terdeteksi (matriks riwayat perjumpaan dengan individu dalam baris dan hari dalam kolom). Kami mengisi setiap baris dengan 1, 2, atau 3 jika individu tersebut terdeteksi berjalan, bersarang, atau tidak terdeteksi, secara berurutan. Pada subset kedua, kami menghitung jumlah total individu yang ditandai yang tandanya teridentifikasi atau tidak teridentifikasi setiap hari. Subset ini digunakan untuk memperhitungkan bias yang dihasilkan dari individu yang ditandai yang ada di orthomosaic tetapi dengan tanda yang tidak teridentifikasi. Pada set data ketiga, kami menyusun jumlah deteksi yang sesuai dengan individu yang unik atau deteksi berulang dari individu yang sama (yaitu hitungan ganda) untuk setiap hari, dengan hanya mempertimbangkan individu yang ditandai yang diklasifikasikan sebagai berjalan. Akhirnya, selain dari data penandaan-penemuan kembali, set data hitungan keseluruhan terdiri dari jumlah total individu yang berjalan dan bersarang yang terdeteksi setiap hari di orthomosaic.
2.3 Pendekatan dan pemasangan model
Dengan menggunakan data pengamatan tanda dan jumlah keseluruhan, kami mengembangkan pendekatan pemodelan baru untuk memperkirakan kelimpahan total populasi agregat sambil memperhitungkan sumber variasi berikut:
- Populasi terbuka : Individu memasuki gundukan pasir untuk pertama kalinya pada hari yang berbeda selama acara bersarang. Individu ini dapat mengunjungi gundukan pasir beberapa kali sebelum bersarang, tetapi mereka tidak kembali ke gundukan pasir setelah bersarang.
- Status individu : Seorang individu dapat berada dalam salah satu dari dua status dalam satu hari: berjalan atau bersarang. Individu yang berjalan biasanya menjelajahi gundukan pasir dan dapat kembali di hari lain, sementara individu yang bersarang biasanya terbenam di pasir dan tidak kembali ke gundukan pasir.
- Tidak tersedianya : Seseorang yang merupakan bagian dari populasi dapat berada di luar gundukan pasir (yaitu di dalam air) selama penerbangan drone dan oleh karena itu tidak akan tersedia untuk dideteksi.
- Hitungan ganda : Beberapa individu yang berjalan selama penerbangan drone dapat muncul lebih dari sekali di orthomosaic.
- Tanda yang tidak teridentifikasi : Mungkin tidak dapat mengidentifikasi beberapa tanda individual karena terhalang pasir.
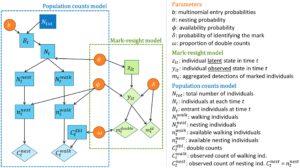
Pendekatan pemodelan yang diusulkan memiliki dua komponen, satu untuk data tanda-penampakan kembali dan satu untuk data jumlah populasi (Gambar 3 ). Kami menyediakan deskripsi model yang lebih terperinci dalam Lampiran S1 . Komponen pertama adalah model penangkapan-penangkapan kembali populasi terbuka multistate untuk data tanda-penampakan kembali (Calvert et al., 2009 ; Kendall et al., 2006 ; White et al., 2010 ) yang diadaptasi untuk menyertakan probabilitas mengidentifikasi tanda individu dan proporsi hitungan ganda (model tanda-penampakan kembali pada Gambar 3 ). Menggunakan formulasi ruang-keadaan (Gimenez et al., 2007 ; Kery & Schaub, 2012 ), kami memodelkan keadaan biologis dan proses deteksi individu setelah penangkapan pertama (yaitu setelah peristiwa penandaan) sebagai hasil kategoris. Proses biologis diatur oleh probabilitas transisi dari keadaan sebenarnya masing-masing pada waktu t ke keadaannya pada waktu t + 1 (Tabel 1a ), sedangkan proses deteksi ditentukan oleh probabilitas mengamati setiap keadaan berdasarkan keadaan sebenarnya (Tabel 1b ).
| (a) Matriks transisi negara | ||||
|---|---|---|---|---|
| Keadaan sebenarnya pada waktu t + 1 | ||||
| Sedang berjalan | Bersarang | Hilang | ||
| Keadaan sebenarnya pada waktu t | Sedang berjalan | (1−θ) | θ | 0 |
| Bersarang | angka 0 | angka 0 | 1 | |
| Hilang | angka 0 | angka 0 | 1 | |
| (b) Matriks deteksi status | ||||
|---|---|---|---|---|
| Keadaan yang diamati | ||||
| Sedang berjalan | Bersarang | Tidak terdeteksi | ||
| Keadaan sebenarnya | Sedang berjalan | ϕjδ | angka 0 | (1−ϕj)+ϕj(1−δ) |
| Bersarang | angka 0 | ϕjδ | (1−ϕj)+ϕj(1−δ) | |
| Hilang | angka 0 | angka 0 | 1 | |


Proporsi individu yang berjalan yang terdeteksi yang sesuai dengan hitungan ganda ditentukan oleh probabilitas hitungan ganda
Akhirnya, kami menggunakan prior yang tidak jelas untuk semua parameter (lihat detailnya di Lampiran S1 ).
Kami menilai identifikasi parameter di bawah struktur model ini menggunakan eksperimen simulasi (lihat tutorial di Lampiran S2 ). Kami melakukan analisis menggunakan pendekatan dua langkah di bawah kerangka kerja Bayesian, dan model dipasang dengan paket Nimble (de Valpine et al., 2017 , 2024 ) di R (Tim Inti R, 2023 ). Kami pertama-tama memperkirakan parameter untuk data mark-resight dan kemudian menggunakan sampel posterior acak dari perkiraan ini untuk memodelkan jumlah keseluruhan (lihat detail di Lampiran S1 ). Kami menilai konvergensi model dengan inspeksi visual traceplot dan menggunakan statistik R-hat (Brooks & Gelman, 1998 ).
3 HASIL
Jumlah kura-kura secara keseluruhan dalam orthomosaic harian bervariasi antara 531 (373 individu berjalan, 158 bersarang) dan 4073 (1934 berjalan, 2139 bersarang), menghasilkan total 26.532 deteksi kura-kura sungai dalam 12 hari (Gambar 5a ). Dari 1187 individu yang ditandai sepanjang 12 hari ini, 468 ditangkap kembali setidaknya satu kali, 61 lebih dari dua kali, dan hanya 7 kura-kura yang terlihat kembali lebih dari tiga kali. Sebanyak 325 (69,4%) dari 468 penampakan kembali individu yang ditandai terjadi pada kesempatan pertama setelah penandaan (yaitu individu ditandai pada malam hari [3 pagi] dan terlihat kembali saat matahari terbit [6 pagi] pada hari yang sama). Proporsi individu yang ditandai yang terdeteksi di setiap hari yang memiliki tanda yang dapat diidentifikasi bervariasi dari 63,6% hingga 87,2%. Akhirnya, jika hanya mempertimbangkan individu yang ditandai dengan tanda pengenal yang diklasifikasikan sebagai berjalan, proporsi hitungan ganda bervariasi dari 6,6% hingga 31,6%.
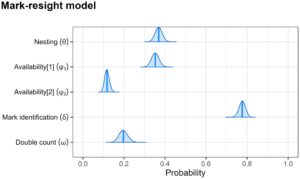
Mengenai parameter yang diestimasikan dalam komponen pemodelan mark-resight (Gambar 4 ), kami menemukan bahwa probabilitas individu untuk bersarang pada setiap kesempatan adalah 0,369 (95% CI = 0,335–0,404). Probabilitas individu yang hadir pada malam hari untuk tersedia dalam penerbangan pukul 6 pagi (
) diperkirakan sebesar 0,353 (95% CI = 0,318–0,389). Di sisi lain, individu yang ditandai dan belum bersarang dalam 1 hari memiliki kemungkinan
sebesar 0,118 (95% CI = 0,096–0,141) yang tersedia pada hari berikutnya. Perkiraan proporsi hitungan ganda untuk individu yang berjalan (
) adalah 0,197 (95% CI = 0,157–0,241) sedangkan probabilitas mengidentifikasi simbol individu yang ditandai (
) adalah 0,777 (95% CI = 0,745–0,807).

Bahasa Indonesia: Selain perbandingan antara model yang berbeda untuk hitungan ortomosaik, kami juga membandingkan hasil kami dengan hitungan tanah visual. Hitungan tanah visual dilakukan secara bersamaan dengan penerbangan drone (6 pagi) oleh tiga pengamat independen yang berlokasi di titik pandang tertinggi gundukan pasir. Jumlah rata-rata per hari bervariasi dari 656 hingga 2.257, berjumlah 15.955 individu selama 12 hari survei. Dibandingkan dengan hitungan ortomosaik, hitungan tanah secara umum lebih rendah daripada jumlah drone total, kecuali pada hari dengan jumlah kura-kura paling sedikit di gundukan pasir (Gambar S1a ). Yang terpenting, perbedaan antara kedua metode meningkat dengan kelimpahan kura-kura yang diperkirakan (Gambar S1b ), menunjukkan bahwa jumlah individu yang lebih banyak di gundukan pasir sering kali menyebabkan lebih banyak halangan dari pandangan permukaan tanah, menghalangi hitungan permukaan tanah yang lebih akurat.
4 DISKUSI
Kami mengembangkan model untuk memperkirakan kelimpahan populasi yang dikumpulkan secara spasial dari hitungan berbasis drone di orthomosaik yang memperhitungkan berbagai sumber kesalahan negatif palsu dan positif palsu serta dinamika temporal individu yang memasuki dan meninggalkan area target. Sejauh pengetahuan kami, ini adalah studi pertama yang memperhitungkan berbagai sumber bias ini secara bersamaan dalam hitungan orthomosaik survei berbasis drone. Pendekatan kami bergantung pada kumpulan data tingkat individu (data penandaan-penglihatan ulang) untuk memperkirakan ketersediaan dan hitungan ganda di bawah model tangkap-tangkap kembali multistatus populasi terbuka. Yang penting, pendekatan populasi terbuka dengan formulasi superpopulasi memungkinkan mengakomodasi dinamika temporal individu dan dengan demikian estimasi kelimpahan total sepanjang seluruh periode studi. Dengan menggunakan contoh penyu sungai selama acara bersarang massal, kami menunjukkan bagaimana kesalahan ini dapat menjadi signifikan dan tidak boleh diabaikan saat menghitung individu di orthomosaik. Misalnya, kami menemukan bahwa hanya 35% individu yang menggunakan gundukan pasir pada malam hari hadir pada saat penerbangan drone. Yang terpenting, kami juga menemukan bahwa rata-rata 20% kura-kura yang terdeteksi berjalan di orthomosaic sesuai dengan hitungan ganda, dan bahwa satu individu dapat muncul hingga tujuh kali di mosaik. Pendekatan kami juga memperhitungkan fakta bahwa beberapa individu yang ditandai yang terdeteksi di orthomosaic memiliki tanda yang tidak dapat diidentifikasi (sekitar 20% untuk kura-kura sungai), yang dapat menyebabkan estimasi parameter yang bias. Dengan membandingkan hasil kami dengan hasil dari pendekatan pemodelan yang jauh lebih sederhana, kami menyoroti pentingnya mempertimbangkan semua sumber variasi ini dalam hitungan, yang menunjukkan bahwa tidak mengakomodasinya dengan benar dapat menyebabkan bias substansial dalam estimasi kelimpahan.
Selama pengembangan model, kami mengidentifikasi beberapa rekomendasi umum untuk merancang survei drone ortomosaik guna menghitung populasi satwa liar yang dikumpulkan secara spasial. Pertama, jika tujuannya adalah untuk memperkirakan jumlah total individu yang menggunakan lokasi pengumpulan sepanjang musim, penting bahwa jendela waktu pengambilan sampel mencakup seluruh periode saat lokasi tersebut digunakan oleh spesies satwa liar sasaran. Agar dapat dilaksanakan, ini mungkin memerlukan pelaksanaan survei pada hari-hari alternatif. Namun, perlu dicatat bahwa jika substitusi populasi diperkirakan tinggi untuk interval waktu yang ditentukan (misalnya terlalu banyak kedatangan dan keberangkatan), menambah interval waktu antara survei dapat menjadi masalah. Lebih jauh, menandai sebagian individu sebelum setiap penerbangan drone (seperti yang kami lakukan) dapat memberikan informasi yang lebih baik tentang variabilitas temporal parameter dibandingkan dengan menandai hanya sekali sebelum memulai survei. Namun demikian, karena individu yang ditandai pada kesempatan awal memiliki riwayat pertemuan yang lebih lama dan dengan demikian dapat memberikan lebih banyak informasi untuk estimasi parameter, seseorang dapat memprioritaskan menandai lebih banyak individu dalam kunjungan pertama ini. Penelitian masa depan yang menggunakan eksperimen simulasi untuk mengevaluasi strategi desain pengambilan sampel, termasuk total durasi survei, interval waktu antara kesempatan, dan kapan menandai individu dapat menjadi penting untuk pengoptimalan survei dalam program pemantauan.
Perumusan model yang kami usulkan menyerupai pendekatan sebelumnya yang menggabungkan data hitungan dan tanda-penglihatan ulang (dengan pita/cincin) untuk memodelkan kelimpahan dengan dinamika temporal di lokasi persinggahan migrasi burung (Lyons et al., 2016 ; Matechou et al., 2013 ; Tucker et al., 2023 ). Namun, pendekatan sebelumnya ini tidak mencakup beberapa status, kemungkinan penghitungan ganda individu, dan keberadaan tanda yang tidak dapat diidentifikasi. Yang penting, Matechou et al. ( 2013 ) mengeksplorasi pengaruh penghitungan ganda dan tanda yang tidak dapat diidentifikasi menggunakan eksperimen simulasi, dan menyimpulkan bahwa tidak memperhitungkan sumber kesalahan ini dapat mengakibatkan estimasi kelimpahan yang berlebihan. Sumber kesalahan lain yang dapat diakomodasi dalam kerangka kerja pemodelan kami adalah kemungkinan salah mendeteksi individu yang bersarang sebagai berjalan, untuk memperhitungkan individu yang muncul dalam berjalan orthomosaic sebelum atau setelah bersarang. Kami secara singkat mengeksplorasi dengan simulasi versi model yang mengakomodasi kesalahan deteksi kondisi individu dengan memasukkan probabilitas kesalahan deteksi tertentu dalam matriks deteksi. Hasil model ini menjanjikan, terutama untuk skenario di mana beberapa individu ditangkap kembali pada beberapa kesempatan setelah penandaan. Namun, kami tidak mempertimbangkan formulasi model ini untuk data kura-kura karena probabilitas kesalahan deteksi diperkirakan sangat mendekati nol, yang menunjukkan bahwa jenis kesalahan deteksi ini tidak signifikan untuk kumpulan data ini.
Pendekatan yang dikembangkan dapat diaplikasikan pada konteks lain di mana populasi yang teragregasi secara spasial disurvei menggunakan orthomosaik yang berasal dari drone. Misalnya, drone telah digunakan untuk mensurvei kura-kura air tawar di area berjemur (Bogolin et al., 2021 ) dan kura-kura laut di lokasi bersarang (Rees et al., 2018 ; Thorson et al., 2012 ). Lebih jauh lagi, orthomosaik adalah pendekatan umum yang digunakan dalam survei lokasi tempat berkumpul dan koloni bersarang atau berkembang biak anjing laut dan burung (misalnya, Goebel et al., 2015 ; Kellenberger et al., 2021 ; Korczak-Abshire et al., 2019 ; Procksch et al., 2023 ; Weinstein et al., 2022 ). Jelas, konteks-konteks yang berbeda ini mungkin memerlukan beberapa adaptasi, seperti pendekatan lain untuk menandai individu. Misalnya, anjing laut ditandai untuk survei drone dengan cara memotong bulunya (Sorrell et al., 2019 ), rusa besar dipasangi kerah visibilitas tinggi untuk dapat dilihat kembali dalam survei udara (Bear et al., 1989 ), dan berbagai spesies ungulata telah ditandai dengan bola cat untuk pengamatan kembali udara (Pauley & Crenshaw, 2006 ; Skalski et al., 2005 ). Adaptasi penting lainnya mengacu pada status individu mana yang akan direpresentasikan. Ketika tidak ada perbedaan yang diharapkan dalam dinamika temporal di antara individu-individu dari kelas yang berbeda, kerangka kerja pemodelan kami dapat disederhanakan untuk hanya mewakili dua status: ada dan pergi. Namun, beberapa populasi agregat dapat menunjukkan perilaku temporal yang berbeda antara jantan dan betina dewasa (dan mungkin juvenil) (misalnya, Dujon et al., 2021 ; Infantes et al., 2022 ), yang berpotensi memerlukan penggunaan jenis kelamin dan/atau usia sebagai beberapa status. Beberapa status individu mungkin juga perlu diperhitungkan ketika mempelajari koloni bersarang burung, di mana tahap sarang (misalnya, pembangunan sarang, inkubasi telur, periode bersarang) dapat memengaruhi dinamika temporal burung dewasa (Gallego & Sarasola, 2021 ; Lachman et al., 2020 ; Sardà-Palomera et al., 2017 ). Akhirnya, penting untuk berpikir hati-hati apakah asumsi populasi terbuka atau populasi tertutup harus dipertimbangkan. Misalnya, ketika mensurvei populasi di mana individu yang sama menggunakan area agregasi selama periode pengambilan sampel (yaitu, entri dan keberangkatan tidak signifikan), model dapat disederhanakan menjadi model tangkap-tangkap kembali populasi tertutup, yang hanya memperkirakan ketersediaan, hitungan ganda, dan identifikasi tanda. Misalnya, anjing laut dewasa dapat menggunakan tempat keluar untuk beristirahat di antara periode makan selama beberapa minggu (Cordes & Thompson, 2015 ), dengan individu yang sama menggunakan area tersebut selama periode ini.
Tipe data lain pada level individu (berbeda dari data mark-resight) juga dapat digunakan dalam kerangka kerja pemodelan yang diusulkan. Misalnya, data pelacakan GPS dari perangkat telemetri telah digunakan untuk memperkirakan kesalahan deteksi dalam survei udara, khususnya untuk mengatasi ketersediaan dan persepsi individu (Barker, 2008 ). Dalam konteks pendekatan kami, jika beberapa individu dilacak telemetri, akan mungkin untuk memperkirakan ketersediaan mereka selama penerbangan drone berdasarkan lokasi mereka. Lebih jauh lagi, dengan menggunakan jejak pergerakan ini selama penerbangan, mungkin saja untuk memodelkan pola pergerakan dan mengidentifikasi (atau memperkirakan) hitungan ganda. Kesimpulannya, pendekatan yang dikembangkan menyediakan kerangka kerja fleksibel yang dapat disesuaikan untuk berbagai spesies dan konteks menurut sifat populasi agregat (yaitu, tertutup atau terbuka), berbagai kemungkinan status individu, dan berbagai tipe data level individu yang akan dikumpulkan.
4.1 Implikasi konservasi dan pengelolaan
Kura-kura Sungai Raksasa Amerika Selatan telah mengalami penurunan historis. Awalnya, kelimpahannya jauh lebih besar, dan bersarang massal terjadi di banyak anak sungai Amazon dan Sungai Orinoco, tetapi baru-baru ini ia telah menghilang dari banyak sungai ini atau sekarang hadir dalam kepadatan yang jauh lebih rendah. Namun, masih ada beberapa populasi besar spesies di seluruh jangkauannya, dan beberapa di antaranya tampaknya pulih (Forero-Medina et al., 2021 ). Perilaku musiman spesies ini, berkumpul dan bersarang di gundukan pasir selama musim kemarau, memberikan peluang yang sangat berharga untuk memantau populasinya. Salah satu metode tradisional untuk memperkirakan jumlah betina yang bersarang adalah menghitung sarang, terutama untuk agregasi kecil. Namun, memperkirakan kelimpahan di daerah dengan bersarang massal yang substansial menggunakan metode seperti itu menjadi menantang atau bahkan mustahil karena sarang individu tidak dapat dibedakan satu sama lain. Metode umum lainnya untuk memperkirakan kelimpahan kura-kura sungai adalah menghitung tukik saat mereka muncul. Namun, menghitung telur penyu yang baru menetas menghadirkan tantangan penting, yang berpotensi memberikan estimasi kelimpahan yang bias (Norris, 2025 ), selain menjadi pendekatan yang lebih invasif dan melelahkan. Yang penting, kami telah menunjukkan bahwa penghitungan visual di tanah juga dapat menjadi pendekatan yang tidak efektif untuk menilai kelimpahan selama peristiwa bersarang massal karena terhalangnya pandangan di permukaan tanah yang disebabkan oleh sejumlah besar individu.
Oleh karena itu, pendekatan yang disajikan memiliki keuntungan penting untuk memantau populasi kura-kura secara agregat. Pertama, citra udara memberikan sudut pandang yang bagus untuk menghitung kura-kura tanpa halangan. Kedua, metode standar yang dapat diterapkan dan dibandingkan di berbagai lokasi dan tahun yang berbeda, dengan estimasi ketidakpastian terkait, memberikan penilaian yang lebih kuat tentang ukuran dan tren populasi. Lebih jauh, menggunakan teknik yang kurang invasif yang mengurangi manipulasi dan gangguan terhadap hewan, seperti survei berbasis drone, sangat penting untuk spesies yang terancam punah. Oleh karena itu, kami memperkirakan pembentukan jaringan kolaboratif lembaga pemerintah dan nonpemerintah di seluruh area bersarang massal kura-kura sungai untuk memantau spesies tersebut menggunakan protokol yang mirip dengan yang kami kembangkan.
Jumlah betina yang diperkirakan dalam penelitian ini menegaskan betapa pentingnya lokasi ini bagi konservasi spesies tersebut. Total kelimpahan yang kami perkirakan untuk lokasi agregasi selama 12 hari bersarang massal berturut-turut adalah 41.377 ekor kura-kura. Perkiraan ini lebih tinggi daripada bersarang massal lainnya yang tercatat untuk spesies tersebut (Forero-Medina et al., 2021 ). Karena peristiwa bersarang berlanjut selama beberapa hari setelah penerbangan pesawat nirawak terakhir, kami dapat berasumsi bahwa beberapa ribu betina tambahan masih menggunakan gundukan pasir khusus ini. Meskipun perkiraan kelimpahan ini mungkin mewakili sejumlah besar kura-kura sungai, itu mungkin hanya sebagian kecil dari populasi historis di wilayah Amazon, berdasarkan catatan historis telur yang diekspor (Forero-Medina et al., 2021 ). Penerapan protokol pemantauan harus mempertimbangkan perluasan survei ke seluruh periode bersarang. Lebih jauh lagi, karena ada gundukan pasir lain di wilayah tersebut yang juga digunakan kura-kura untuk bersarang, penting untuk memasukkannya guna memperoleh perkiraan komprehensif populasi bersarang di wilayah tersebut.
Agregasi musiman populasi satwa liar (misalnya, lokasi pengangkutan, lokasi persinggahan migrasi, koloni bersarang atau berkembang biak) memberikan peluang besar untuk memperkirakan dan memantau kelimpahan secara efisien. Hitungan berbasis drone telah digunakan untuk mensurvei populasi agregat spasial (Christie et al., 2016 ; Lyons et al., 2019 ), tetapi ada sedikit kesadaran tentang beberapa kesalahan deteksi yang dapat memengaruhi jumlah dan akibatnya bias estimasi populasi. Penggunaan orthomosaik yang dihasilkan dari penerbangan drone menjadi pendekatan yang semakin umum untuk mensurvei populasi satwa liar agregat dan, karena alasan ini, kami percaya metodologi yang dikembangkan memiliki potensi besar untuk diterapkan (dan diadaptasi) ke banyak konteks berbeda di mana spesies yang terancam disurvei menggunakan orthomosaik berbasis drone. Pada akhirnya, kami berharap bahwa pendekatan ini akan berkontribusi pada pemantauan kelimpahan yang efisien dan tepat waktu dalam program konservasi dan pengelolaan satwa liar.



Tinggalkan Balasan