
Abstrak
- Jalan merupakan infrastruktur yang ada di mana-mana yang memiliki dampak buruk terhadap satwa liar dan berkontribusi terhadap peningkatan kematian dan fragmentasi populasi hewan. Meskipun beberapa langkah mitigasi tersedia untuk mengurangi dampak jalan, perencanaannya jarang mempertimbangkan sifat lingkungan yang dinamis, yang dapat tercermin dalam variasi temporal dalam kesesuaian habitat dan konektivitas bagi spesies hewan. Akibatnya, efektivitas langkah-langkah tersebut mungkin tidak memenuhi harapan.
- Dengan menggabungkan citra satelit beresolusi tinggi dan pemodelan konektivitas, kami mengusulkan pendekatan yang dapat digeneralisasi untuk mengidentifikasi titik persimpangan yang paling memungkinkan di suatu penghalang pada waktu yang berbeda. Informasi ini mungkin penting dalam merencanakan langkah-langkah mitigasi yang dapat memperhitungkan komponen dinamis.
- Kami mengumpulkan data kejadian tiga anuran yang beradaptasi dengan lahan pertanian dan citra multispektral Sentinel-2 definisi spasiotemporal tinggi untuk membangun model kesesuaian habitat yang menangkap hubungan antara lingkungan dan spesies dalam area studi (area pertanian yang dilintasi jalan raya). Kami kemudian memproyeksikan model tersebut ke enam potret dari dua musim kawin amfibi berikutnya. Terakhir, kami menggunakan model konektivitas berbasis teori sirkuit untuk mengidentifikasi area dengan probabilitas tertinggi untuk dilintasi jalan raya untuk setiap potret/spesies.
- Kami menemukan perbedaan yang mencolok dari waktu ke waktu untuk setiap spesies, baik dalam hal kesesuaian maupun konektivitas. Lebih jauh, distribusi dan kepentingan relatif titik-titik persimpangan berubah drastis antara 2 tahun dan juga dalam kurun waktu yang sama. Beberapa titik persimpangan stabil dari waktu ke waktu, sementara yang lain penting untuk gambaran tertentu.
- Sintesis dan aplikasi . Tidak mempertimbangkan variabilitas spasiotemporal lingkungan dapat menyebabkan hilangnya informasi penting saat memodelkan lokasi tempat aliran hewan yang diprediksi berpotongan dengan jalan raya. Dinamika ekosistem yang terdampak jalan dapat diperhitungkan menggunakan data penginderaan jarak jauh yang tersedia secara bebas. Ini dapat menjadi elemen penting dalam mencapai tujuan menjaga konektivitas dan meminimalkan kematian satwa liar.
1. PENDAHULUAN
Jalan sering kali merupakan kejahatan yang diperlukan yang biasanya muncul bersamaan dengan pembangunan ekonomi suatu wilayah dan merupakan ancaman besar bagi keanekaragaman hayati, misalnya, memengaruhi kepadatan spesies dan mengganggu aliran ekologi horizontal (Bennett, 2017 ; Fahrig & Rytwinski, 2009 ; Forman & Alexander, 1998 ). Fitur lanskap yang ada di mana-mana ini memiliki efek buruk langsung dan tidak langsung pada satwa liar dan ekosistem. Sebagai dampak langsung, kita dapat menyebutkan peningkatan kematian hewan akibat tabrakan kendaraan, hilangnya habitat dan degradasi, serta polusi (termasuk kebisingan). Di sisi lain, dampak tidak langsung terutama terkait dengan fragmentasi habitat. Kombinasi dari efek-efek ini menghasilkan infrastruktur yang pada dasarnya mengurangi konektivitas habitat melalui pembatasan pergerakan satwa liar (Benítez-López et al., 2010 ; Bennett, 2017 ; Forman & Alexander, 1998 ; Hamer et al., 2021 ; Herrera-Montes & Aide, 2011 ; Jochimsen et al., 2004 ; Tulloch et al., 2019 ; van der Ree et al., 2015 ). Konektivitas lanskap, memang, dapat didefinisikan sebagai ‘sejauh mana lanskap memfasilitasi atau menghambat pergerakan di antara petak-petak sumber daya’ (Taylor et al., 1993 ). Konektivitas yang berkurang mengubah aliran gen dan dinamika metapopulasi dan dapat memiliki efek merugikan pada ketahanan populasi hewan, berinteraksi dengan peningkatan tingkat kematian (Borda-de-Água et al., 2011 ; Epps et al., 2005 ; Fahrig et al., 1995 ; Roger et al., 2011 ).
Fakta yang membuat dampak negatif infrastruktur semakin nyata dan mengancam adalah bahwa jaringan jalan global terus tumbuh, dan diperkirakan akan terus tumbuh dalam beberapa dekade mendatang. Pertumbuhan ini semakin menonjol di negara-negara berkembang, dalam beberapa kasus mengancam beberapa wilayah dengan keanekaragaman hayati tertinggi di planet ini (Gallice et al., 2019 ; Hughes, 2019 ; Prasniewski et al., 2020 ).
Karena beratnya masalah ini, kebutuhan untuk menemukan jalan tengah dan mematuhi kawasan lindung dan konvensi internasional masih terbuka. Mitigasi dapat menjadi alat penting untuk mengurangi dampak buruk jalan terhadap keanekaragaman hayati. Beberapa langkah mitigasi telah diusulkan dan diterapkan untuk mengurangi angka kematian dan meningkatkan konektivitas antara petak habitat melalui, misalnya, jalan layang, pagar, dan terowongan (Glista et al., 2009 ). Struktur ini dapat secara efektif berkontribusi pada konservasi satwa liar tetapi memerlukan proyeksi yang memadai (Hamer et al., 2023 ; Lesbarrères & Fahrig, 2012 ).
Dalam perspektif ini, model konektivitas telah menjadi alat penting dan tersebar luas untuk perencanaan dan penentuan prioritas konservasi (Kučas et al., 2023 ; Lechner et al., 2015 ; Loro et al., 2015 ; Mimet et al., 2016 ; Paemelaere et al., 2023 ). Model-model ini terutama didasarkan pada ‘permukaan resistansi’, yaitu, lapisan raster, yang mewakili bentang alam, di mana nilai resistansi ditetapkan untuk setiap piksel. Nilai ini bergantung pada ekologi spesies target dan biasanya mempertimbangkan jenis habitat, risiko kematian, dan setiap faktor lain yang dapat memengaruhi biaya pergerakan melalui sel (Kumar & Cushman, 2022 ; Zeller et al., 2012 ). Peta resistansi kemudian dianalisis dengan algoritma, yang menyediakan estimasi konektivitas (Adriaensen et al., 2003 ; Bunn et al., 2000 ; McRae et al., 2008 ).
Bentang alam adalah sistem yang dinamis, yang dapat berubah seiring waktu, baik karena penyebab alamiah maupun tekanan yang disebabkan oleh manusia (Turner et al., 1993 ; Zhou et al., 2018 ). Variasi intratahunan dan intertahunan pada properti bentang alam seperti fenologi dan hidrologi, serta pada tahap-tahap siklus hidup organisme (misalnya reproduksi/hibernasi atau juvenil/dewasa) memengaruhi kesesuaian habitat pada skala temporal yang berbeda (Brambilla et al., 2012 ; Gonçalves et al., 2016 ; Hill et al., 2015 ; Medinas et al., 2021 ). Perubahan-perubahan ini semakin diperhitungkan saat memodelkan konektivitas ekologis, yang mengarah pada konsep konektivitas dinamis (Zeller et al., 2020 ). Studi terbaru telah menyoroti bagaimana evaluasi konektivitas ‘snapshot’ tidak memberikan hasil yang dapat diandalkan di area dengan variabel spasiotemporal tinggi seperti yang memiliki ketersediaan air permukaan yang tidak konstan (Bishop-Taylor et al., 2018 ; Simpkins, 2017 ). Oleh karena itu, ketika merencanakan langkah-langkah mitigasi penghalang, dinamika lanskap, yang tercermin dalam fluktuasi konektivitas ekologis, harus dipertimbangkan. Ini akan memastikan bahwa tindakan kompensasi memberikan permeabilitas yang efektif sepanjang tahun atau selama fase terpenting dari siklus biologis untuk spesies fokus. Untuk mengejar tujuan seperti itu, menjadi penting untuk memodelkan konektivitas lanskap secara akurat, termasuk variasinya baik dalam waktu maupun ruang.
Dalam studi ini, kami menyajikan kasus nyata dari perencanaan mitigasi jalan di lanskap pertanian, yang dicirikan oleh variabilitas kondisi lingkungan antartahunan (rotasi tanaman) dan intratahunan (fase budidaya, ketersediaan air). Dengan berfokus pada amfibi dan bentangan jalan raya yang dibatasi, tujuan kami adalah untuk mengidentifikasi lokasi dengan probabilitas lintasan tertinggi dan variasi dalam distribusi dan kepentingannya. Untuk ini, kami secara eksplisit mempertimbangkan variabilitas temporal lingkungan sekitar karena menentukan konektivitas lanskap khusus spesies. Kami memilih pendekatan yang sangat didorong oleh data dengan menggunakan titik kejadian yang dikumpulkan secara ad hoc dan citra multispektral resolusi tinggi, yang disediakan oleh misi ESA Sentinel-2 (Martimort et al., 2007 ). Data mentah ini digunakan untuk membangun model kesesuaian habitat (HSM) dan model konektivitas berbasis teori sirkuit (McRae et al., 2008 ).
Kami menggunakan amfibi sebagai taksa model karena dua alasan utama: (1) mereka sangat rentan terhadap fragmentasi habitat dan kematian di jalan raya karena karakteristik ekologisnya, karena mereka membentuk metapopulasi, bergerak lambat dan membutuhkan tipe habitat yang berbeda selama siklus hidupnya (Beebee, 2013 ; Elzanowski et al., 2009 ; Fahrig et al., 1995 ; Lehtinen et al., 1999 ); (2) mereka biasanya menunjukkan pola pergerakan musiman karena ketergantungan mereka pada air untuk reproduksi; akibatnya, kita dapat memperkirakan variabilitas spasiotemporal dalam kondisi lingkungan akan memiliki efek nyata pada konektivitas fungsional mereka (Duellman & Trueb, 1986 ; Sinsch, 1990 ).
Tujuan akhir dari makalah ini adalah untuk menggarisbawahi pentingnya mempertimbangkan perubahan spasiotemporal dalam konektivitas saat merencanakan penyeberangan jalan dan tindakan mitigasi serupa. Kami mengusulkan pendekatan yang dapat diterapkan secara luas, yang mudah dipindahkan ke taksa dan situasi lain yang tunduk pada dinamika tersebut.
2 BAHAN DAN METODE
2.1 Lokasi penelitian
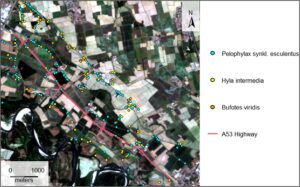
Daerah studi (Gambar 1 ) adalah zona pertanian antara kota Pavia dan Bereguardo, yang terletak di Dataran Po (Lombardy, Italia). Daerah tersebut membentang lebih dari 26,5 km 2 dan dilintasi oleh jalan raya A53, sepanjang 9,1 km. Jalan raya tersebut tidak memiliki jalur satwa liar, tetapi memiliki terowongan terutama untuk tujuan drainase, yang, dengan tindakan yang tepat, dapat dimanfaatkan oleh hewan kecil. Jenis penutup lahan sebagian besar adalah tanaman, dengan kehadiran besar rumpun poplar, sedangkan habitat alami (hutan dataran rendah residual dan daerah aliran sungai) langka dan terkonsentrasi di sepanjang Sungai Ticino. Dalam konteks ini, habitat utama yang digunakan oleh amfibi untuk reproduksi adalah yang terkait dengan kegiatan pertanian, seperti kanal, parit, dan sawah. Karakteristik yang dijelaskan adalah umum untuk lahan pertanian dataran Po, salah satu daerah dataran rendah terbesar di Eropa selatan. Dataran Po juga merupakan salah satu wilayah yang paling berkembang di semua sektor ekonomi dan, sebagai konsekuensinya, sangat dipengaruhi oleh polusi, urbanisasi, konsumsi lahan dan fragmentasi habitat (Romano & Zullo, 2016 ). Baru-baru ini, wilayah tersebut mengalami perubahan drastis dalam praktik budaya dengan efek langsung pada badan air yang terkait dengan pertanian: memang, distrik padi Italia semakin mengadopsi praktik penyemaian kering , yang melibatkan penundaan banjir, mengurangi waktu ketersediaan air permukaan (Ranghetti & Boschetti, 2022 ) dan dengan efek buruk pada keanekaragaman hayati, misalnya, pada Ardeidae (Fasola et al., 2022 ). Iklim wilayah tersebut adalah kontinental dengan rezim curah hujan kontinental subalpin. Khususnya, tahun kedua dari dua tahun pengambilan sampel (yaitu 2022) ditandai dengan kekeringan yang parah (Montanari et al., 2023 ), khususnya pada fase awal periode aktivitas spesies target dan pada bulan-bulan sebelumnya (lihat Tabel S1 ). Kombinasi variabilitas yang berkaitan dengan rotasi tanaman dan pertumbuhan perkebunan serta perubahan ketersediaan air akibat banjir di sawah dan curah hujan membuat area dan periode studi optimal untuk mewakili lingkungan dinamis yang harus dihadapi.
2.2 Spesies sasaran
Sebagai spesies target, kami menggunakan tiga anuran ( Bufotes viridis , Hyla intermedia dan Pelophylax synkl. esculentus ), yang mewakili totalitas amfibi yang tersebar luas di wilayah studi (Dataran Po, Italia) (Sindaco et al., 2006 ). Spesies ini memang beradaptasi dengan baik terhadap lingkungan pertanian di Italia utara. Mereka adalah spesies dataran rendah yang khas, dengan valensi ekologis yang besar, dan mampu mengeksploitasi berbagai jenis habitat untuk berkembang biak, termasuk badan air yang terganggu dan buatan. Lebih jauh lagi, mereka sedikit berbeda dalam ekologi dan fenologi mereka. B. viridis adalah spesies pionir yang bereproduksi di perairan dangkal dan seringkali sementara dari Maret hingga Juli. Hyla intermedia dapat berkembang biak dari Maret hingga Agustus di berbagai habitat, mungkin dikelilingi oleh tepi bervegetasi, tempat orang dewasa dapat memanjat. Akhirnya, Pelophylax synkl. esculentus biasanya berkembang biak dari April hingga Juli, lebih disukai di lokasi dengan air permanen, stagnan atau mengalir lemah (Lanza et al., 2007 ).
2.3 Alur kerja umum
Kami menggunakan lima langkah utama untuk mengidentifikasi titik dan area penyeberangan jalan yang paling memungkinkan (Gambar 2 ): (1) mengumpulkan data kejadian tentang spesies target sepanjang musim reproduksi (yaitu tempat pergerakan dan aktivitas keseluruhan lebih tinggi); (2) memproses citra Sentinel-2 dan mengekstrak metrik berbasis penginderaan jauh; (3) membangun model kesesuaian habitat (HSM) yang menangkap hubungan kejadian-lingkungan untuk setiap spesies; (4) memproyeksikan HSM ke dalam snapshot spesifik; (5) memperoleh model konektivitas dengan menggunakan kesesuaian habitat yang diproyeksikan dan penghalang yang ada untuk mengidentifikasi simpul lokal dan mengatur peta resistensi spesifik waktu; (6) untuk setiap snapshot dan untuk setiap spesies target, mengidentifikasi titik jepit dalam model konektivitas yang mewakili titik penyeberangan yang paling mungkin.
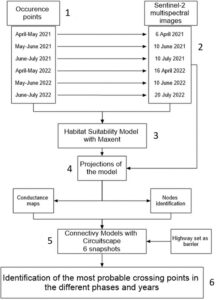
2.3.1 Pengumpulan data kejadian (1)
Data dikumpulkan dari bulan April hingga Juli tahun 2021 dan 2022, selama puncak aktivitas untuk tiga spesies target (Lanza et al., 2007 ).
Upaya pengambilan sampel konsisten selama periode yang dipertimbangkan. Kami secara sistematis mensurvei 32 lokasi, yang tersebar di area studi (lihat Gambar S1 ). Di sana, kami mendengarkan panggilan amfibi selama 5 menit dan secara visual menjelajahi sawah dan parit di sekitarnya. Setiap lokasi disurvei sekitar setiap 15–20 hari dengan total 4–6 pengambilan sampel per tahun. Survei dilakukan pada malam hari, mulai dari matahari terbenam dan seterusnya, selama sekitar 3 jam. Sebanyak 35 survei dilakukan: 20 pada tahun 2021, 15 pada tahun 2022. Kami mengintegrasikan data kejadian dengan lokasi amfibi yang terlihat atau terdengar saat berpindah di antara lokasi pengambilan sampel. Karena metode survei tidak memerlukan penangkapan atau penanganan hewan dan tidak dapat membahayakan mereka dengan cara apa pun, baik persetujuan etis maupun izin khusus tidak perlu diperoleh.
2.3.2 Ekstraksi metrik berbasis penginderaan jauh (2), pemodelan kesesuaian habitat (3) dan proyeksi (4)
Untuk memperoleh HSM resolusi tinggi, berdasarkan kondisi lingkungan yang sedekat mungkin dengan kondisi yang terjadi pada saat spesies terdeteksi, kami menggunakan citra satelit Sentinel-2 sebagai sumber prediktor lingkungan.
Dalam studi ini, kami menggunakan produk Level-2A Sentinel-2, yang menyediakan citra Reflektansi Permukaan yang dikoreksi untuk hamburan dan penyerapan yang terjadi di atmosfer (Panduan Pengguna—Sentinel-2 MSI—Pemrosesan Level-2—Sentinel Online, 2024 ). Kami mengunduh tiga citra multispektral untuk setiap tahun pengambilan sampel data, menghindari citra dengan tutupan awan. Kami memilih tiga snapshot setahun, yang mencakup fase-fase utama periode studi tahunan dan aktivitas amfibi, untuk menangkap perbedaan dalam kondisi lingkungan, tergantung pada pertumbuhan vegetasi, banjir sawah, dan ketersediaan air secara umum: (i) April: fase budidaya awal dengan beberapa ladang masih dibajak dan banjir sawah hampir tidak ada. Awal perkembangbiakan amfibi di mana air tersedia. (ii) Juni: banjir sawah. Puncak aktivitas perkembangbiakan untuk spesies target. (iii) Juli: fase budidaya akhir, pertumbuhan vegetasi maksimum. Fase menurun dari aktivitas perkembangbiakan amfibi (terutama B. viridis ).
Kami menggunakan semua 12 pita spektral (SB) yang disediakan oleh produk Level-2A sebagai prediktor, tanpa interpretasi apriori, yang memungkinkan model mengidentifikasi pita dan kombinasi yang relevan untuk memprediksi distribusi spesies (Sacchi et al., 2022 ). Gambar dipotong di area studi, dan resolusi peta diseragamkan menjadi 20 m. Dengan demikian, hanya dua pita yang memerlukan penurunan skala spasial, yang tersedia pada resolusi 60 m. Kami memilih resolusi berbutir halus untuk menangkap elemen lanskap kecil yang penting bagi spesies target, seperti kanal dan sebagian kecil sawah yang tergenang air.
Karena (i) tujuan HSM adalah untuk memodelkan hubungan ekologis antara kejadian dan variabel lingkungan (yaitu nilai SB), dan (ii) nilai SB berubah selama periode studi, kami mengaitkan titik kejadian dengan nilai SB terdekat dalam waktu. Titik yang dikumpulkan pada bulan April dan awal Mei dikaitkan dengan nilai SB dari peta satelit April, titik pada akhir Mei dan awal Juni dari peta Juni, dan titik pada akhir Juni dan Juli dari peta Juli. Kami mengulangi prosedur tersebut selama 2 tahun. Dengan menggabungkan semua data untuk setiap spesies, kami memperoleh satu set data komprehensif yang mampu menangkap seluruh informasi yang tersedia tentang hubungan spesies-lingkungan dan siap untuk pembangunan model. Pendekatan ini dimaksudkan untuk menghasilkan model distribusi spesies yang dinamis, menghindari ketidaksesuaian antara lokasi spesies dan variabel prediktor (Milanesi et al., 2020 ). Model distribusi spesies yang dinamis telah terbukti mengatasi model statis, terutama ketika diterapkan pada area yang mengalami variabilitas lingkungan yang kuat (Ingenloff & Peterson, 2021 ; Srivastava & Carroll, 2023 ).
Kami menerapkan pendekatan entropi maksimum yang banyak digunakan untuk pemodelan kesesuaian habitat dari data kehadiran saja, yang diimplementasikan dalam perangkat lunak Maxent 3.4.4 (Phillips et al., 2006 ), dengan menggunakan paket R dismo (Hijmans et al., 2011 ; R Core Team, 2022 ). Pendekatan ini telah terbukti berkinerja lebih baik daripada sebagian besar algoritme lain (misalnya Random Forest, Support Vector Machine), terutama dalam pemodelan dengan data kehadiran saja (Valavi et al., 2022 ). Untuk mengurangi kompleksitas model dan risiko overfitting, kami tidak menggunakan parameter Maxent default tetapi hanya menggunakan transformasi linier dan kuadrat dari variabel prediktor (Low et al., 2021 ). Kami mengambil sampel secara acak 2000 titik latar belakang di area studi yang nilai SB-nya diekstraksi untuk masing-masing dari enam snapshot. Jadi, kita memiliki 2000 titik dalam pengertian spasial, tetapi 12.000 (2000 × 6) titik untuk membangun model. Kumpulan titik latar belakang sama untuk semua spesies.
Model-model tersebut divalidasi menggunakan prosedur pelipatan silang: kumpulan data kejadian dibagi secara acak menjadi lima subset dengan ukuran yang sama, masing-masing dengan distribusi sampel kehadiran/latar belakang yang sama; selanjutnya, empat subset digunakan untuk menghasilkan model, dan yang terakhir digunakan untuk pengujian. Performa model diestimasi menggunakan AUC (area under the receiver operating Characteristics curve), yang merepresentasikan ukuran kemampuan model untuk membedakan antara lokasi kehadiran dan latar belakang yang tidak bergantung pada ambang batas (Liu et al., 2005 ; Merow et al., 2013 ). Namun, AUC tidak mempertimbangkan autokorelasi spasial karena prosedur validasi silang (yaitu bias dari urutan spasial; Segurado et al., 2006 ; Veloz, 2009 ) dan pengelompokan geografis titik-titik kehadiran (Hijmans, 2012 ). Oleh karena itu, prosedur yang diusulkan oleh Hijmans ( 2012 ) juga digunakan untuk menghitung AUC terkoreksi yang memperhitungkan autokorelasi spasial (pwdAUC).
Seluruh prosedur dijawab untuk masing-masing dari tiga spesies target, menghasilkan tiga model terpisah. Kami memproyeksikan tiga HSM akhir ke setiap citra multispektral snapshot untuk memperoleh peta kesesuaian waktu tertentu untuk setiap spesies target (6 × 3 = 18 proyeksi HSM).
2.3.3 Pemodelan konektivitas (5) dan identifikasi titik persimpangan (6)
Kami memodelkan konektivitas dengan menerapkan pendekatan sukses yang diusulkan oleh McRae dkk. ( 2008 ), berdasarkan teori sirkuit klasik. Dalam metode ini, bentang alam direpresentasikan sebagai permukaan konduktif dengan resistansi variabel, sementara lokasi kejadian atau area yang sesuai adalah simpul tempat arus masuk atau keluar; dengan memecahkan sirkuit, intensitas arus dihitung di setiap sel raster. Nilai intensitas ini dapat diinterpretasikan sebagai aliran hewan yang diprediksi melalui sel.
Di sini, untuk setiap snapshot, kami menggunakan estimasi kesesuaian yang disediakan oleh proyeksi HSM untuk memperoleh peta konduktansi dan menetapkan sebagai node saat ini area dengan kesesuaian di atas ambang batas spesifik spesies (Spesifisitas Maksimum plus Sensitivitas; Liu et al., 2013 . P. synkl. esculentus: 0,52, H. intermedia : 0,48, B. viridis : 0,58) dan dengan perluasan setidaknya 1000 m 2 . Kami memodelkan jalan raya sebagai penghalang, memberinya nilai konduktansi rendah (ditetapkan pada 200 pada skala 0–1000). Analisis dilakukan menggunakan perangkat lunak Circuitscape 5 dalam bahasa pemrograman Julia (Anantharaman et al., 2019 ; Bezanson et al., 2017 ).
Karena tujuannya bukanlah untuk memperkirakan konektivitas area secara umum, tetapi untuk mengidentifikasi titik persimpangan yang paling mungkin melalui jalan raya, kami menjalankan Circuitscape dalam mode lanjutan , dengan menetapkan semua simpul di satu sisi jalan raya sebagai simpul sumber , dan semua simpul di sisi lain sebagai simpul dasar . Operasi diulang dalam arah yang berlawanan untuk memodelkan aliran di kedua arah, dan kedua model dijumlahkan dan diskalakan antara 0 (tidak ada arus) dan 1 (arus maksimum). Analisis dilakukan untuk masing-masing dari tiga spesies target, dan untuk keenam snapshot yang digunakan untuk pemodelan kesesuaian habitat, untuk mendapatkan rute pergerakan utama dan titik persimpangan yang paling mungkin pada enam momen berbeda dari periode pengambilan sampel 2 tahun.
Sebagai pendekatan pertama, sebelum melakukan penskalaan, kami mengukur variasi temporal intensitas arus di area sekitar jalan raya dengan membuat zona penyangga sejauh 1000 m di sekitar jalan raya dan membandingkan variasi intensitas arus di dalamnya pada enam potret.
Bahasa Indonesia: Untuk mendeteksi area penyeberangan yang paling memungkinkan di sepanjang jalan raya dari keluaran model konektivitas untuk setiap spesies/gambaran potret, pertama-tama kami mendefinisikan sebagai titik penyeberangan (CP) kelompok dari sedikitnya 4 sel arus tinggi yang berdekatan (yaitu intensitas arus relatif >0,7; area ≥1600 m 2 ) yang memotong jalan raya. Kami kemudian menggabungkan CP dari enam gambar potret dalam satu lapisan unik, dan kami mengelompokkannya menggunakan analisis klaster (metode UPGMA; Sokal & Michener, 1958 ), menetapkan jarak antar klaster ke 500 m. Kami menghitung convex hull untuk setiap klaster untuk mengidentifikasi area dengan probabilitas tinggi untuk dilewati. Akhirnya, kami memotong area ini dengan CP dari setiap gambar potret. Dengan cara ini, kami secara otomatis mengidentifikasi area dengan keberadaan CP yang terus-menerus di seluruh spesies dan gambar potret. Kami membandingkan dan mengevaluasi hasil dari pendekatan yang baru saja dijelaskan dengan memeriksa peta konektivitas secara visual.
Semua analisis spasial dilakukan dengan paket R terra (Hijmans, 2023 ).
3 HASIL
3.1 Pengumpulan data kejadian
Sebanyak 793 titik keberadaan dikumpulkan selama dua tahun pengambilan sampel ( P. synkl. esculentus : 466; H. intermedia : 190; B. viridis : 137; Gambar 1 ). Titik-titik tersebut dikurangi menjadi 570 ( P. synkl. esculentus : 302; H. intermedia : 153; B. viridis : 115) setelah menskalakan kejadian ke resolusi spasial variabel lingkungan yang sama. Tabel yang menunjukkan perincian berdasarkan cuplikan disediakan dalam Informasi Pendukung (lihat Tabel S2 ).
3.2 Pemodelan kesesuaian habitat
Ketiga HSM menghasilkan nilai AUC berikut: P. synkl esculentus : 0,67 ± 0,02 SD; H. intermedia : 0,73 ± 0,05 SD; B. viridis : 0,71 ± 0,03 SD. Nilai pwdAUC (lihat Tabel S3 ) serupa atau bahkan lebih besar (min. 0,70; maks. 0,80), yang tidak mencerminkan efek struktur spasial variabel lingkungan (yaitu pita spektral Sentinel-2) pada kemampuan model untuk membedakan titik keberadaan dan latar belakang.
Enam dari dua belas Pita Spektral yang digunakan sebagai prediktor menunjukkan pentingnya permutasi yang relevan untuk ketiga spesies: B04, B08/8A, B01, B03, B12, B06 (Tabel S4 ; Gambar 2 ).
Proyeksi HSM pada enam citra multispektral Sentinel-2, yang sesuai dengan fase awal, tengah, dan akhir musim kawin amfibi pada tahun 2021 dan 2022, menunjukkan pola variasi kesesuaian yang serupa. Khususnya: (1) ketiga spesies tersebut dipengaruhi oleh perubahan dalam distribusi area yang sesuai; (2) nilai kesesuaian relatif lebih tinggi pada cuplikan pusat (Juni) di kedua tahun pengambilan sampel; (3) rata-rata, kesesuaian habitat 21% lebih rendah (April: 25% lebih rendah, Juni: 13% lebih rendah, Juli: 26% lebih rendah) pada tahun 2022 dibandingkan dengan tahun 2021.
B. viridis menunjukkan nilai kesesuaian yang jauh lebih tinggi baik pada cuplikan awal maupun cuplikan tengah dibandingkan dengan cuplikan tengah. Selain itu, pada cuplikan tengah, kesesuaian lebih terkonsentrasi, dengan beberapa petak memiliki nilai yang sangat tinggi (Gambar 3 ).
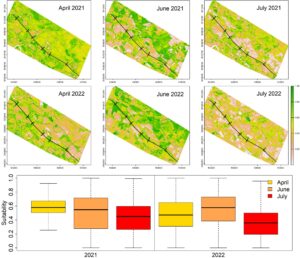
Untuk P. synkl. esculentus pada tahun 2021 nilai kesesuaian cukup stabil dari waktu ke waktu. Akan tetapi, distribusi area yang sesuai berubah, dengan bagian utara jalan raya khususnya sesuai pada potret tengah. Pada tahun 2022, kesesuaian rendah pada potret pertama dan kemudian naik ke nilai yang sebanding dengan tahun pertama (lihat Gambar S3 ).
H. intermedia memiliki tingkat kesesuaian yang rendah pada gambaran awal namun tetap mempertahankan nilai yang tinggi pada bulan Juni dan Juli; selain itu, spesies ini merupakan spesies yang mengalami penurunan kesesuaian paling parah pada tahun 2022 (lihat Gambar S4 ).
Secara umum, perubahan kesesuaian lahan mengikuti perubahan kondisi lingkungan di wilayah tersebut, yaitu pertumbuhan vegetasi dan terutama keberadaan air di kanal dan persawahan. Perbandingan antara proyeksi kesesuaian habitat dengan citra True Color Sentinel-2 menunjukkan bahwa wilayah dengan kesesuaian tinggi terutama terkait dengan persawahan dan kanal yang tergenang (Gambar S6 ).
3.3 Model konektivitas dan titik penyeberangan
Analisis konektivitas memberikan gambaran yang jelas tentang area dengan kemungkinan tertinggi untuk dilalui oleh spesies target, dengan perhatian khusus pada mereka yang melintasi jalan raya, yang sangat penting dalam penelitian ini (Gambar 4 ; Gambar S6 dan S7 ).

Intensitas arus di sekitar jalan raya mengalami variasi yang nyata di antara snapshot yang dipertimbangkan, secara umum lebih rendah pada tahun 2022 dibandingkan pada tahun 2021 dan mengikuti tren musiman yang berbeda di antara ketiga spesies (Gambar 5 ). H. intermedia menunjukkan nilai intensitas rendah pada bulan April, dengan peningkatan pada bulan Juni yang bertahan hingga akhir musim. B. viridis memiliki nilai intensitas arus yang sama dalam dua snapshot pertama tahun 2021, menurun pada yang terakhir. Sebaliknya, pada tahun 2022, ia memiliki puncak intensitas arus dalam snapshot tengah, dengan nilai yang lebih rendah baik sebelum maupun sesudahnya. Tren musiman dalam intensitas arus dalam 2 tahun juga berbeda untuk P. synkl. esculentus : pada tahun 2021, intensitas arus cukup stabil, sedangkan pada tahun 2022, snapshot pertama memiliki nilai yang jauh lebih rendah.
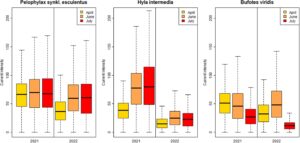
Untuk setiap spesies di setiap snapshot yang dipertimbangkan, adalah mungkin untuk mengidentifikasi satu atau lebih titik penyeberangan, dengan variasi penting dalam distribusi spasial mereka (ratusan meter) dan jumlah (1 hingga 18), baik antar tahun dan intraannual (Gambar 6 ). Secara keseluruhan, total 7 area dengan probabilitas tinggi untuk lintasan telah diidentifikasi (Gambar 7 ). Hanya satu dari mereka yang ditemukan mencakup CP untuk semua spesies di semua snapshot; terletak di dekat pintu keluar ketiga jalan raya dari utara. Tidak ada area lain selain ini yang berisi CP untuk satu spesies dalam snapshot apa pun. Namun, yang lain mendukung tiga spesies dalam satu atau lebih snapshot.





Tinggalkan Balasan