Keranjang Anda kosong!
Penyaringan dan analisis mekanistik varietas kedelai yang tahan terhadap sindrom stay-green yang disebabkan oleh virus terkait stay-green kedelai

Kedelai, salah satu tanaman pangan paling penting secara ekonomi di dunia, menyumbang 25% dari produksi minyak nabati global dan menyediakan dua pertiga protein pekat untuk pakan ternak. Namun, dalam beberapa tahun terakhir, munculnya sindrom tetap hijau kedelai (SGS), atau ‘zhengqing’, di wilayah Huang-Huai-Hai di Tiongkok, telah menjadi ancaman besar bagi produksi kedelai. Sindrom ini telah menyebabkan kerugian hasil panen yang parah, dengan beberapa daerah melaporkan gagal panen total (Li et al ., 2019 ; Wei et al ., 2023 ; Xu et al ., 2019 ). Masalah ini meluas dan telah menjadi perhatian utama di seluruh Tiongkok (Cheng et al ., 2023 ; He et al ., 2025 ; Xu et al ., 2019 ).
SGS ditandai dengan penuaan yang tertunda saat panen, dengan daun kedelai yang tetap hijau, polong gagal terbentuk, dan perkembangan biji terhenti, yang menyebabkan polong sedikit atau datar (Cheng et al ., 2022 ; Li et al ., 2019 ; Wang et al ., 2022 ; Wei et al ., 2023 ). Dampak mendalam dari sindrom ini pada hasil dan kualitas menghadirkan tantangan yang signifikan bagi industri kedelai di Tiongkok. Baru-baru ini, penelitian kami telah mengidentifikasi etiologi SGS sebagai geminivirus baru, yang disebut soybean stay-green-associated geminivirus (SoSGV). Melalui postulat Koch, kami telah menunjukkan untuk pertama kalinya bahwa SoSGV menyebabkan gejala stay-green pada kedelai (Cheng et al ., 2022 ). SoSGV adalah virus DNA untai tunggal monopartit, yang kemungkinan besar dihasilkan dari rekombinasi intergenerik geminivirus (Cheng et al ., 2022 ). Lebih jauh, kami telah mengidentifikasi wereng coklat biasa ( Orosius orientalis ) sebagai vektor penularan SoSGV (Cheng et al ., 2023 ). Laporan sebelumnya telah mengindikasikan bahwa SGS menghadirkan beragam ekspresi fenotip di lapangan, dengan variasi dalam tingkat keparahan gejala (Cheng et al ., 2023 ; Xu et al ., 2019 ). Akan tetapi, penyebab variabilitas ini tidak jelas, dan tidak pasti apakah perbedaan dalam ekspresi gejala terkait dengan infeksi SoSGV di berbagai kultivar kedelai.
Pemuliaan tanaman inang yang tahan merupakan strategi yang hemat biaya dan ramah lingkungan untuk meningkatkan hasil panen. Akan tetapi, kemajuan dalam mengidentifikasi varietas kedelai yang tahan terhadap SoSGV masih terbatas. Dalam penelitian ini, kami menggunakan sistem inokulasi yang ditularkan melalui vektor yang dikembangkan di laboratorium kami untuk menilai ketahanan terhadap SoSGV pada 36 plasma nutfah kedelai unggul dari wilayah Huang-Huai-Hai (Tabel S1 ). Temuan kami mengungkapkan bahwa sebagian besar varietas ini sangat rentan terhadap SoSGV.
Untuk menentukan apakah SoSGV menyebabkan gejala tetap hijau saat matang, enam belas varietas kedelai yang terinfeksi SoSGV dan kontrol tiruan dipindahkan ke ladang satu bulan setelah inokulasi. Varietas-varietas ini, yang dipilih karena fenotipenya yang relatif lemah selama infeksi SoSGV tahap awal, dipantau dari Juli hingga Oktober (2022–2024). Semua tanaman yang diberi perlakuan tiruan memperlihatkan biji yang menguning dan matang, sementara 15 dari 16 varietas, kecuali Fudou_18, memperlihatkan penuaan daun yang tertunda dan polong yang pipih (Gambar 1a,b ). Sepuluh varietas (Jiadou_2, Jidou_12, Xiangfeng_4, Andou_203, Huadou_19, Lindou_11, Xudou_20, Shengdi_1, Lindou_10, dan Zhonghuang_13) memperlihatkan pengerdilan saat matang. Lima varietas yang tersisa (Wandou_38, Tianyikedou_18, Hedou_12, Huaidou_11, dan Shidou_1) tidak menunjukkan perubahan signifikan pada tinggi tanaman (Gambar 1a ; Gambar S2a ). Jumlah polong berkurang pada sebagian besar varietas kerdil, kecuali Xudou_20 (Gambar 1b ; Gambar S2b ). Pada Xudou_20 dan Shidou_1, jumlah polong tidak terpengaruh, tetapi pengisian polong sangat berkurang, dengan lebih dari 80% polong datar (Gambar S2c ). Hasil ini menunjukkan bahwa variasi fenotipik di antara varietas kedelai terwujud setelah infeksi SoSGV.
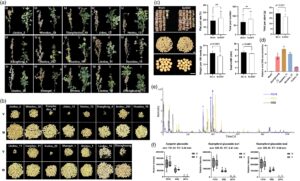
Untuk menilai dampak infeksi SoSGV pada hasil, kami menganalisis varietas kedelai yang terinfeksi dan mengamati kehilangan hasil berkisar antara 15,8% hingga 97,05% (Gambar 1b,c ; Gambar S3a ). Di antara varietas ini, sepuluh menunjukkan pengurangan hasil yang parah melebihi 80%, termasuk Jiadou_2, Wandou_38, Tianyikedou_18, Jidou_12, Xiangfeng_4, Huadou_19, Lindou_11, Xudou_20, Lindou_10, dan Zhonghuang_13 (Gambar S3a ). Menariknya, Huaidou_11 menunjukkan kehilangan hasil terendah (36,19%) meskipun menunjukkan gejala tetap hijau yang nyata (Gambar 1a ). Kehilangan hasil yang relatif rendah ini sejalan dengan pengamatan kami bahwa infeksi SoSGV tidak secara signifikan mempengaruhi jumlah polong total atau tingkat polong datar di Huaidou_11 (Gambar S3b ). Namun, Huaidou_11 yang terinfeksi menghasilkan lebih banyak benih abnormal, ditandai dengan ukuran yang lebih kecil, bentuk yang berubah, dan pematangan yang tidak sempurna, sehingga sangat mengurangi kualitas benih (Gambar 1b ; Gambar S3b ).
Fudou_18 menunjukkan toleransi yang luar biasa terhadap infeksi SoSGV, seperti yang tercermin dalam tingkat keparahan gejala yang ringan (Gambar 1c ; Gambar S1 ). Sementara tanaman menunjukkan gejala seperti daun melengkung dan kerdil selama tahap awal infeksi (Gambar S1 ), hanya gejala tetap hijau ringan yang diamati saat dewasa, tanpa perbedaan signifikan dalam tinggi tanaman atau pematangan polong dibandingkan dengan perlakuan tiruan (Gambar 1c , Gambar S1b ). Infeksi SoSGV tidak secara signifikan mempengaruhi jumlah polong total atau tingkat polong datar di Fudou_18 (Gambar 1c ). Meskipun ada sedikit pengurangan lebar biji rata-rata dari 7,705 mm menjadi 6,872 mm (Gambar 1c ), berat 100 butir hanya berkurang 18%. Total hasil per tanaman berkurang sekitar 15,8% – dampak yang relatif kecil dibandingkan dengan varietas lain yang mengalami kehilangan hasil lebih besar (Gambar 1c ). Selain itu, benih Fudou_18 yang terinfeksi SoSGV mempertahankan bentuk yang seragam, konsisten dengan benih kontrol (Gambar 1c ).
Untuk menyelidiki apakah gejala ringan yang diamati pada Fudou_18 dikaitkan dengan titer virus yang lebih rendah, kami melakukan reaksi berantai polimerase kuantitatif menggunakan daun yang terinfeksi untuk mengukur akumulasi SoSGV pada tahap R5 di Zhonghuang_13, Huaidou_11, Wandou_38, dan Fudou_18. Hasilnya menunjukkan bahwa akumulasi SoSGV paling rendah pada Fudou_18 dibandingkan dengan tiga kultivar lainnya (Gambar 1d ), yang menunjukkan bahwa Fudou_18 menunjukkan resistensi terhadap infeksi SoSGV, kemungkinan melalui mekanisme yang membatasi akumulasi virus.
Berikutnya, untuk menjelaskan mekanisme molekuler yang mendasari resistensi Fudou_18 terhadap SoSGV, kami melakukan analisis transkriptomik dan metabolomik pada Fudou_18 dan dua kultivar yang rentan (Williams 82 dan Zhonghuang 13) setelah infeksi SoSGV. Analisis transkriptom pertama kali mengungkap gen yang diekspresikan secara diferensial (DEG) antara tanaman yang terinfeksi versus tanaman yang diobati secara semu (Gambar S4a ; Tabel S2 ) dengan analisis KEGG yang mengungkap bahwa gen metabolik (terutama untuk metabolisme khusus) diperkaya secara signifikan (Gambar S4b ). Kami selanjutnya melakukan analisis metabolomik non-target pada sampel yang sama Fudou_18, Williams 82, dan Zhonghuang 13 yang digunakan untuk analisis transkriptom.
Analisis komponen utama (PCA) menunjukkan pemisahan yang jelas dalam profil metabolik global di antara kultivar (Gambar S4c ). Sementara Fudou_18 yang terinfeksi SoSGV dan kultivar yang rentan hanya menunjukkan perubahan metabolom minor dibandingkan dengan kontrol (Gambar S4c ). Analisis metabolit yang terakumulasi secara diferensial (DAM) mengungkapkan bahwa ada diferensiasi yang jelas dari profil metabolik di antara ketiga kultivar dan sejumlah besar DAM meningkat secara signifikan di Fudou_18 dibandingkan dengan kultivar yang rentan (Gambar S4d ; Tabel S3 ). Analisis jalur pengayaan DAG mengidentifikasi bahwa jalur KEGG yang diperkaya secara signifikan tersebut terlibat dalam metabolisme fenilpropanoid dan pati dan sukrosa di Fudou_18 pasca infeksi, yang menunjukkan bahwa fenolik dan gula mungkin mengalami pemrograman ulang metabolik setelah infeksi SoSGV (Gambar S4e,f ). Kami selanjutnya melakukan analisis perbandingan antara Fudou_18 dan dua kultivar yang rentan dengan memetakan perbedaan yang signifikan versus perubahan lipatan dari setiap fitur massa (Gambar 1e ). Khususnya, kaempferol glukosida ( m/z 595,16 @6,4 dan 6,7 menit, berturut-turut) dan apigenin glukosida ( m/z 741,22 @5,4 menit) sangat terakumulasi di Fudou_18, lebih sedikit terakumulasi di Williams 82 dan hanya terakumulasi di Zhonghuang 13 (Gambar 1e,f ), suatu pola yang sangat konsisten dengan ketahanan SoSGV dari ketiga kultivar.
Untuk memvalidasi peran jalur flavonoid/isoflavonoid dalam resistensi Fudou_18 terhadap SoSGV, chalcone synthase 1 ( CHS1 , Glyma.19G105100 dan Glyma.06G118550), enzim kunci dalam jalur ini, dibungkam menggunakan apple latent spherical virus-induced gene silencing (VIGS). Membungkam ekspresi CHS1 dalam Fudou_18 secara signifikan mengkompromikan resistensi tanaman terhadap SoSGV, sebagaimana dibuktikan oleh peningkatan akumulasi DNA SoSGV dan protein lapisan (CP), bersama dengan pengerdilan tanaman yang parah pada tahap awal dan akhir infeksi (Gambar S5 ). Selain itu, membungkam ekspresi CHS1 juga meningkatkan laju polong datar setelah infeksi SoSGV dibandingkan dengan kontrol (Gambar S5f ). Kami juga menggunakan galur kedelai yang secara stabil mengekspresikan isoflavone synthase 2 ( IFS2 ) kedelai, enzim kunci yang terlibat dalam akumulasi isoflavone (Jung et al ., 2000 ). Setelah inokulasi SoSGV, ekspresi berlebihan IFS2 pada kultivar kedelai ‘Jack’ secara signifikan meningkatkan ketahanan tanaman terhadap SoSGV dibandingkan dengan tanaman kontrol ‘Jack’, sebagaimana dibuktikan oleh penurunan akumulasi DNA SoSGV dan CP (Gambar S6 ). Temuan ini menyoroti peran penting jalur flavonoid/isoflavonoid dalam ketahanan kedelai terhadap SoSGV.
Secara kolektif, melalui penyaringan skala besar, Fudou_18 sebagai varietas yang toleran terhadap SoSGV. Mengungkap mekanisme ketahanannya memberikan dasar yang menjanjikan untuk mengembangkan kultivar kedelai dengan ketahanan yang lebih baik terhadap penyakit virus yang kritis ini.



Tinggalkan Balasan