
Abstrak
Waterhemp ( Amaranthus tuberculatus ) adalah gulma berdaun lebar yang penting secara ekonomi yang mengancam produksi jagung dan kedelai di seluruh Amerika Serikat. Biotipe waterhemp (CT_Res [biotipe resistan dari Connecticut]) yang bertahan dari beberapa aplikasi glifosat diidentifikasi dari ladang jagung di Connecticut (CT). Studi rumah kaca dan laboratorium dilakukan untuk (1) mengonfirmasi resistensi glifosat pada biotipe waterhemp CT_Res dan (2) menyelidiki apakah resistensi glifosat pada biotipe CT_Res disebabkan oleh mekanisme berbasis target-situs. Studi dosis-respons menunjukkan bahwa biotipe CT_Res 5,8 kali lipat lebih resistan terhadap glifosat dibandingkan dengan biotipe rentan yang diketahui (NE_Sus) dari Nebraska. Tidak ada mutasi titik yang terdeteksi pada posisi Pro 102 atau Thr 106 pada gen EPSPS dari biotipe CT_Res. Uji reaksi berantai polimerase kuantitatif mengungkapkan bahwa salah satu dari tiga tanaman waterhemp CT_Res memiliki jumlah salinan gen EPSPS 3,5 kali lebih tinggi (dibandingkan dengan gen CPS housekeeping ), sedangkan dua tanaman lainnya tidak menunjukkan amplifikasi gen EPSPS . Jelas, amplifikasi gen EPSPS sebagian menjelaskan resistensi glifosat pada biotipe waterhemp resistan glifosat yang baru diidentifikasi dari CT, yang menunjukkan bahwa mekanisme alternatif mungkin ada. Penelitian ini melaporkan kasus pertama resistensi glifosat dan amplifikasi gen EPSPS pada waterhemp dari Connecticut dan menyoroti perlunya penerapan strategi pengendalian gulma yang beragam untuk mencegah penyebarannya lebih lanjut.
Ringkasan Bahasa Sederhana
Waterhemp adalah salah satu spesies gulma yang paling merepotkan di Midwest AS. Dalam beberapa tahun terakhir, biotipe waterhemp juga diidentifikasi di Amerika Serikat bagian timur laut, termasuk Connecticut dan New York. Penelitian ini bertujuan untuk mengonfirmasi keberadaan resistensi glifosat dalam biotipe waterhemp yang dikumpulkan dari ladang jagung di Connecticut dan menyelidiki apakah resistensi glifosat disebabkan oleh mekanisme berbasis target-situs. Studi rumah kaca menyimpulkan bahwa biotipe waterhemp dari Connecticut 5,8 kali lebih tahan terhadap glifosat jika dibandingkan dengan biotipe rentan glifosat yang diketahui dari Nebraska. Lebih jauh lagi, satu dari tiga tanaman waterhemp yang tahan glifosat menunjukkan salinan gen target ( EPSPS ) 3,5 kali lebih tinggi. Secara keseluruhan, hasil ini mengonfirmasi laporan pertama resistensi glifosat dalam biotipe waterhemp yang baru-baru ini diidentifikasi dari Connecticut. Petani perlu waspada dan harus mengadopsi strategi pengendalian gulma yang beragam untuk mencegah penyebarannya lebih lanjut.
Singkatan
CPS
karbamoil fosfat sintetase
CT
Connecticut
CT_Res
biotipe resistan dari Connecticut
CT_Sus
biotipe rentan dari Connecticut
EPSP
5-enolpiruvilsikimat-3-fosfat sintase
GR
tahan glifosat
NE_Sus
biotipe rentan dari Nebraska
1. PENDAHULUAN
Komersialisasi tanaman tahan glifosat (GR) pada pertengahan 1990-an dan budidayanya yang meluas menyebabkan ketergantungan yang hampir eksklusif pada glifosat untuk pengendalian gulma spektrum luas. Pada tahun 2005, sekitar 45 juta kg glifosat diaplikasikan pada jutaan ha jagung GR ( Zea mays L.), kapas ( Gossypium hirsutum L.), dan kedelai [ Glycine max (L.) Merr.] dibandingkan dengan <6 juta kg pada tahun 1996 (NASS, 1997, 2006 ). Secara historis, glifosat telah menjadi herbisida pascatumbuh yang paling ekonomis dan efektif untuk mengendalikan berbagai spesies gulma pada tanaman GR (Askew & Wilcut, 1999 ; Aulakh et al., 2011 ; Price et al., 2008 ). Namun, penggunaan glifosat yang terus-menerus dan berulang mengakibatkan evolusi biotipe gulma GR. Rumput kuda [ Conyza canadensis (L.) Cronq.] merupakan spesies gulma pertama yang mengembangkan resistensi terhadap glifosat di lahan kedelai GR di Delaware (VanGessel, 2001 ). Sejak saat itu, beberapa spesies gulma lainnya, termasuk waterhemp, telah mengembangkan resistensi terhadap glifosat (Aulakh et al., 2021 ; Kumar et al., 2025 ; Legleiter & Bradley, 2008 ; Price et al., 2011 ).
Waterhemp adalah salah satu spesies gulma yang paling agresif dan mengganggu dalam produksi jagung dan kedelai di Midwest AS (Hager et al., 2002 ; Steckel & Sprague, 2004b ). Persaingan waterhemp dengan tanaman agronomis sering kali mengakibatkan kerugian ekonomi yang signifikan. Misalnya, hasil panen sorgum dan kedelai berkurang hingga 37%–45% ketika waterhemp dibiarkan bersaing selama 10 minggu (Feltner et al., 1969 ; Hager et al., 2002 ; Steckel & Sprague, 2004a ). Demikian pula, gangguan waterhemp sepanjang musim mengurangi hasil panen jagung hingga 11%–74% (Steckel & Sprague, 2004b ).
Waterhemp yang resistan terhadap herbisida merupakan masalah serius di Amerika Serikat (Heap, 2025 ). Waterhemp yang resistan terhadap glifosat pertama kali dilaporkan di Missouri dan kini telah dikonfirmasi di 21 negara bagian di Amerika Serikat dan Ontario di Kanada (Heap, 2025 ). Beberapa biotipe waterhemp yang resistan terhadap herbisida juga telah dilaporkan dari Illinois dan Missouri (Evans et al., 2019 ; Shergill et al., 2018 ). Berbagai mekanisme molekuler dan fisiologis yang memberikan resistensi glifosat pada gulma telah ditemukan (Chatham et al., 2015 ; Dinelli et al., 2008 ; Ge et al., 2010 ; Kumar et al., 2015 ; Perez-Jones et al., 2007 ; Shaner et al., 2011 ). Misalnya, rumput gandum tahunan GR ( Lolium rigidum ), rumput angsa ( Eleusine indica ), dan rumput gandum Italia ( Lolium multiflorum ) memiliki mutasi titik pada gen 5-enolpyruvylshikimate-3-phosphate synthase ( EPPS ) (Perez-Jones et al., 2007 ; Wakelin & Preston, 2006 ; Yu et al., 2015 ). Biotipe tertentu rumput gandum tahunan GR dan rumput kuda menunjukkan penurunan penyerapan dan translokasi glifosat (Dinelli et al., 2008 ; Lorraine-Colwill et al., 2003 ). Ge et al. ( 2010 ) melaporkan peningkatan sekuestrasi vakuolar glifosat dalam biotipe rumput kuda GR. Peneliti lain mengamati amplifikasi gen EPSPS sebagai dasar resistensi glifosat pada kochia ( Bassia scoparia ), Palmer amaranth ( Amaranthus palmeri ), dan waterhemp (Aulakh et al., 2024 ; Chahal et al., 2017 ; Gaines et al., 2010 ; Kumar et al., 2015 ). Alterasi situs target ganda yang melibatkan amplifikasi gen EPSPS dan mutasi titik Pro 106 Ser juga telah didokumentasikan pada GR waterhemp (Nandula et al., 2013 ; Shaner et al., 2011 ). Biotipe GR waterhemp dengan 3–9 salinan gen EPSPS telah dilaporkan dari Illinois (Chatham et al., 2015 ). Di sisi lain, pada GR Palmer amaranth dari Connecticut dan Georgia, lebih dari 100 salinan gen EPSPS hadir (Aulakh et al., 2024 ; Gaines et al., 2010 ).
Selama musim tanam 2023, seorang petani jagung di Connecticut mengamati kegagalan pengendalian gulma waterhemp setelah beberapa kali aplikasi glifosat. Tujuan penelitian ini adalah untuk (1) mengonfirmasi ketahanan terhadap glifosat pada biotipe gulma waterhemp dari Connecticut dan (2) menentukan apakah mekanisme target-situs memberikan ketahanan terhadap glifosat pada biotipe ini.
2 BAHAN DAN METODE
2.1 Bahan tanaman
Sekitar 15 kepala benih dari biotipe GR waterhemp yang diduga bertahan dari dua aplikasi glifosat berurutan pada 840 g ae ha −1 dikumpulkan pada akhir musim panas tahun 2023 dari ladang jagung di Hartford County, CT. Kepala benih dirontokkan secara manual, dan benih dibersihkan secara menyeluruh menggunakan blower kolom udara vertikal dan disimpan dalam kantong polietilen kedap udara pada suhu 5°C hingga digunakan dalam penelitian ini. Biotipe GR waterhemp yang diduga dari Connecticut diberi nama “CT_Res” (biotipe tahan dari Connecticut). Selain itu, benih dari biotipe waterhemp rentan glifosat yang diketahui sebelumnya (NE_Sus) dari ladang dekat Clay Center, Nebraska, digunakan (Kumar et al., 2025 ).
2.2 Percobaan dosis-respons glifosat
Percobaan rumah kaca dilakukan di Connecticut Agricultural Experiment Station, Windsor, CT, dan Guterman Bioclimatic Laboratory di Cornell University di Ithaca, NY, selama musim gugur tahun 2023 dan diulang pada musim semi tahun 2024. Benih biotipe waterhemp CT_Res dan NE_Sus ditanam secara terpisah ke dalam pot plastik persegi (10 × 10 × 10 cm) yang berisi campuran pot komersial (media tanam Pro-Mix Premium All Purpose di Windsor, CT dan campuran Cornell di Ithaca, NY). Pro-Mix Premium All Purpose mengandung gambut sphagnum (80%–90%), humus gambut, perlit, batu kapur, dan teknologi mikoriza PTB297, sedangkan campuran Cornell mengandung gambut, vermikulit, perlit, kapur dolomit, campuran media Jack’s 10-5-10 plus II, dan kalsium sulfat. Bibit waterhemp dikurangi menjadi satu tanaman per pot segera setelah muncul. Bibit diberi air dan nutrisi yang cukup dan disimpan di rumah kaca yang dijaga pada suhu 32/27°C siang/malam dengan periode cahaya 16/8 jam siang/malam yang dilengkapi dengan lampu halida natrium di kedua lokasi. Bibit waterhemp muda (tahap 4 hingga 6 daun dan tinggi 7–10 cm) dari biotipe CT_Res dan NE_Sus diperlakukan dengan glifosat (MADDOG; Loveland Products, Inc., dan Durango, Corteva Agrisciences). Percobaan di kedua lokasi disusun dalam rancangan blok lengkap acak dengan pengaturan faktorial 7 × 2 perlakuan dan 10–15 ulangan (setiap ulangan adalah 1 pot tanaman −1 ). Dua faktor perlakuan adalah (1) tujuh tingkat glifosat (0, 0,5×, 1×, 2×, 4×, 8×, dan 16×), di mana 1× adalah tingkat penggunaan lapangan glifosat (840 g ae ha −1 ) dan (2) dua biotipe waterhemp (CT_Res dan NE_Sus). Perlakuan glifosat disiapkan dalam air deionisasi yang dicampur dengan surfaktan nonionik (Induce; Helena Chemical Co.) pada 250 mL L −1 di Windsor, CT, sedangkan perlakuan glifosat disiapkan dalam air ledeng dan mengandung amonium sulfat (AMS) pada 20 g L −1 di Ithaca, NY. Perlakuan diterapkan dengan penyemprot ransel CO2 terkompresi melalui nosel AI8002VS kipas datar tunggal (TeeJet Technologies) yang dikalibrasi untuk menghasilkan volume semprotan 187 L ha −1 pada 207 kPa dan 3,5 km h −1 di Windsor, CT. Sebaliknya, perlakuan glifosat diterapkan menggunakan ruang semprot kabinet stasioner (Research Track Sprayer, De Vries Manufacturing, RR 1 Box 184) yang dilengkapi dengan nosel kipas datar 8002XR (TeeJet Technologies) yang dikalibrasi untuk menghasilkan 141 L ha −1larutan semprot pada 276 kPa. Tanaman yang diberi perlakuan dikembalikan ke rumah kaca dan disiram setiap hari untuk mempertahankan pertumbuhan yang memadai. Pada 21 hari setelah perlakuan (DAT) di kedua lokasi, tanaman waterhemp dipanen di permukaan tanah dan dikeringkan pada suhu 65°C–72°C selama 5–7 hari untuk menentukan biomassa kering pucuk di atas tanah. Pengurangan biomassa kering pucuk di atas tanah dihitung sebagai persentase kontrol yang tidak diberi perlakuan seperti yang ditunjukkan dalam Persamaan ( 1 ):

2.3 Analisis Statistik
Data mengenai pengurangan biomassa pucuk kering (%) dari biotipe waterhemp CT_Res dan NE_Sus dirata-ratakan di kedua lokasi (dua kali uji coba di setiap lokasi) karena interaksi perlakuan × uji coba × lokasi tidak signifikan ( p = 0,472). Model log-logistik tiga parameter (Persamaan 2 ) dipasang pada pengurangan biomassa pucuk kering menggunakan paket “drc” dalam perangkat lunak R (perangkat lunak statistik R; R Foundation for Statistical Computing) (Knezevic et al., 2007 ):
![]()
di mana Y adalah persentase pengurangan biomassa kering pucuk, x adalah tingkat glifosat, d adalah pengurangan berat kering pucuk maksimum, e adalah nilai GR 50 (jumlah glifosat yang dibutuhkan untuk pengurangan 50% dalam biomassa kering pucuk), dan b merupakan kemiringan relatif di sekitar parameter ” e “. Tingkat resistensi dihitung dengan membagi nilai GR 50 yang diperkirakan dari biotipe yang resistan (CT_Res) dengan biotipe yang rentan (NE_Sus).
2.4 Mekanisme resistensi glifosat
2.4.1 Isolasi DNA genomik
Tanaman GR yang diduga dari Connecticut ditanam menggunakan media tanam dan kondisi rumah kaca yang sama yang sebelumnya disebutkan dalam bioassay respons dosis glifosat. Sebelum perlakuan glifosat, semua tanaman diberi tanda satu per satu, dan jaringan daun segar (100 mg) dikumpulkan secara terpisah dari setiap tanaman. Jaringan daun segar (dipanen sebelum perlakuan glifosat) dari tanaman yang mati pada tingkat glifosat 0,5× digunakan sebagai biotipe sensitif (CT_Sus) (satu tanaman), dan tanaman yang bertahan hidup pada 6720 g ae ha −1 glifosat diberi label sebagai biotipe CT_Res (tiga tanaman). Ekstraksi gDNA dilakukan mengikuti GeneJET Plant Genomic DNA Purification Kit (Thermo Fisher Scientific). Kuantifikasi DNA yang diekstraksi dilakukan dengan NanoDrop One C (Thermo Fisher Scientific).
2.4.2 Mutasi situs target
Wilayah terkonservasi dari gen EPSPS yang mengandung kodon Pro 106 dan Thr 102 diperkuat untuk biotipe waterhemp CT_Res dan CT_Sus melalui reaksi berantai polimerase (PCR). Mutasi situs target EPSPS dievaluasi dengan menargetkan wilayah gen yang sebelumnya diidentifikasi mengandung polimorfisme pada posisi 102 dan 106 yang terkait dengan resistensi terhadap glifosat dalam waterhemp (Nandula et al., 2013 ). Primer (EPSPS Sanger F: 5′-TTGGACGCTCTCAGAACTCTTG GT-3′ dan EPSPS Sanger R: 5′-TGAATTTCCTCCAGCAACGGCAAC-3′) digunakan untuk memperkuat wilayah 192 bp dari ekstraksi DNA genom EPSPS (Nandula et al., 2013 ). PCR dilakukan menggunakan GoTaq Green Master Mix (Promega) dengan parameter berikut: 95°C selama 5 menit diikuti oleh 35 siklus 95°C selama 30 detik, 60°C selama 30 detik, dan 72°C selama 30 detik. Amplikon PCR dibersihkan untuk pengurutan menggunakan GeneJET Gel Extraction and DNA Cleanup Micro Kit (Thermo Fisher Scientific) dan diserahkan untuk pengurutan Sanger dengan primer yang menyertainya di Keck DNA Sequencing Core di Universitas Yale (New Haven, CT). Hasil pengurutan diselaraskan dan dianalisis secara visual menggunakan alat Benchling Alignment (Benchling). Urutan EPSPS dari biotipe CT_Res diselaraskan dengan urutan EPSPS biotipe CT_Sus referensi untuk menentukan substitusi pada kodon Pro 106 atau Thr 102 .
2.4.3 Amplifikasi gen EPSPS
DNA genomik digunakan untuk mengukur jumlah salinan gen EPSPS dalam biotipe CT_Res relatif terhadap gen CPS (gen housekeeping) dengan PCR waktu nyata (Quantum Studio 5, Thermo Fisher) dan protokol Power Track SYBR Green Master Mix (Thermo Fisher). PCR kuantitatif dilakukan menggunakan protokol yang dijelaskan dalam Ma et al. Secara singkat, primer yang menargetkan EPSPS dan gen referensi karbamoil fosfat sintetase ( CPS ) dinormalisasi menjadi 10 µM dan dikombinasikan secara individual dengan 100 ng DNA tanaman dalam reaksi dengan Syber Green PCR Mastermix (Applied Biosystems). Urutan primer bersumber dari Ma et al. ( 2013 ) adalah sebagai berikut: CPS F(5′-ATTGATGCTGCCGAGGATAG-3′), CPS R (5′-GATGCCTCCCTTAGGTTGTTC-3′), EPSPS F (5′-GGTTGTGGTGGTCTGTTTCC-3′), dan EPSPS R (5′-CATCGCTGTTCCTGCATTTC-3′). Setiap sampel biologis dianalisis dalam rangkap tiga menggunakan Sistem QuantStudio 3. Kuantifikasi relatif jumlah salinan EPSPS dilakukan menggunakan metode 2 −ΔΔCt yang dijelaskan oleh Gaines et al. ( 2010 ). Salinan gen EPSPS dalam biotipe CT_Res dinilai relatif terhadap biotipe rentan glifosat yang diketahui (CT_Sus).
3 HASIL DAN PEMBAHASAN
3.1 Resistensi terhadap glifosat
Studi dosis-respons mengonfirmasi bahwa populasi putative waterhemp (CT_Res) yang diidentifikasi dari ladang jagung di Connecticut resistan terhadap glifosat (Tabel 1 ). Berdasarkan model yang disesuaikan, estimasi dosis glifosat yang diperlukan untuk mengurangi bobot kering pucuk CT_Res sebanyak 50% (nilai GR 50 ) adalah 2121 g ha −1 , yang secara signifikan lebih tinggi daripada NE_Sus (363 g ha −1 ). Dibandingkan dengan biotipe NE_Sus, biotipe CT_Res 5,8 kali lipat resistan terhadap glifosat berdasarkan nilai GR 50 (Tabel 1 ; Gambar 1 ). Tingkat resistensi glifosat yang diamati pada biotipe waterhemp CT_Res sebanding dengan tingkat resistensi glifosat (5,6 hingga 9 kali lipat) yang dilaporkan pada GR waterhemp dari Illinois, Iowa, Missouri, New York (Kumar et al., 2025 ; Lorentz et al., 2014 ; Smith & Hallett, 2006 ). Sebaliknya, tingkat resistensi glifosat yang tinggi (17 hingga 39 kali lipat) telah dilaporkan pada populasi GR waterhemp dari Texas, Missouri, dan Nebraska (Legleiter & Bradley, 2008 ; Sarangi et al., 2012 ; Singh et al., 2020 ).
| Estimasi parameter (± SE) a | ||||||
|---|---|---|---|---|---|---|
| Biotipe | D | B | GR 50 | 95% CI | GR 90 | R/S b |
| CT_Res | 94.8 (1.8) | 2.0 (0.1) | tahun 2121 | Tahun 1967–2274 | 6166 | 5.8 |
| NE_Sus | 98.2 (1.4) | 1.4 (0.1) | 363 | 323–402 | tahun 1734 | – |
Singkatan: CI, selang kepercayaan; CT_Res, biotipe waterhemp yang diduga resisten glifosat yang dikumpulkan dari ladang jagung di Hartford County, CT; NE_Sus, biotipe waterhemp yang diketahui rentan terhadap glifosat yang dikumpulkan dari lokasi ladang dekat Clay Center, Nebraska; SE, kesalahan baku. Parameter d merupakan pengurangan biomassa pucuk maksimum, b merupakan kemiringan setiap kurva, dan GR50 merupakan dosis efektif (g ha−1 ) glifosat yang dibutuhkan untuk pengurangan berat kering pucuk sebesar 50% (% yang tidak diobati) untuk setiap biotipe waterhemp. b R/S merupakan perbandingan nilai GR 50 dari biotipe yang diduga resisten terhadap glifosat dengan nilai GR 50 dari biotipe yang rentan.
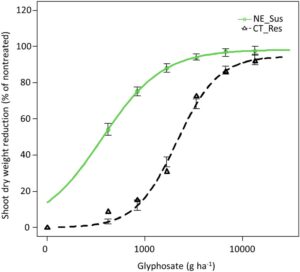
Tingkat glifosat yang diperkirakan untuk mengurangi berat kering pucuk biotipe CT_Res hingga 90% (nilai GR 90 ) adalah 6166 g ha −1 dibandingkan dengan 1734 g ha −1 untuk biotipe NE_Sus. Biotipe waterhemp CT_Res 3,5 kali lipat lebih tahan terhadap glifosat dibandingkan dengan biotipe NE_Sus berdasarkan nilai GR 90 (Tabel 1 ; Gambar 1 ).
3.2 Mekanisme resistensi glifosat
3.2.1 Mutasi target-situs
Mutasi non-sinonim yang menyebabkan perubahan asam amino pada posisi 102 (Thr 102 ) dan/atau 106 (Pro 106 ) pada gen EPSPS sebelumnya telah dilaporkan memberikan resistensi terhadap glifosat (García et al. 2020 ). Analisis sekuens gen EPSPS dari biotipe CT_Res dan CT_Sus tidak mengungkapkan mutasi pada kodon 102 dan 106 (Gambar 2 ). Hasil penelitian ini konsisten dengan Shergill et al. ( 2018 ), yang sebelumnya tidak melaporkan adanya mutasi titik target-situs yang terkait dengan resistensi terhadap glifosat pada populasi waterhemp resisten enam arah dari Missouri. Sebaliknya, Nandula et al. ( 2013 ) mendokumentasikan mutasi target-situs (substitusi asam amino prolin menjadi serin pada posisi 106) dan pengurangan translokasi glifosat pada populasi waterhemp GR dari Mississippi.

3.2.2 Amplifikasi gen EPSPS
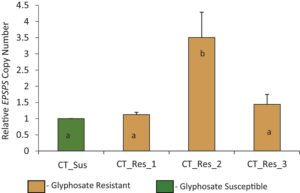
Toksisitas glifosat pada tanaman disebabkan oleh kapasitasnya untuk secara kompetitif menghambat aktivitas EPSPS pada tanaman dan amplifikasi gen EPSPS adalah mekanisme resistensi glifosat yang paling sering diamati (Chatham et al., 2015 ). Hasil PCR kuantitatif mengungkapkan bahwa salah satu tanaman yang diuji dari biotipe CT_Res memiliki salinan gen EPSPS 3,5 kali lipat lebih tinggi dibandingkan dengan CT_Sus (satu salinan) (Gambar 3 ). Namun, dua tanaman lain yang diuji dari biotipe CT_Res mengungkapkan satu salinan gen EPSPS , yang menunjukkan kemungkinan mekanisme alternatif resistensi glifosat (Gambar 3 ). Shergill et al. ( 2018 ) melaporkan lima kali lipat lebih banyak salinan gen EPSPS dalam populasi waterhemp yang resistan terhadap glifosat dan lima MOA herbisida lain dari Missouri. Demikian pula, Dillon et al. ( 2017 ) mendokumentasikan 2–4, 4–8, dan 8–16 salinan gen EPSPS pada tanaman yang sedikit, sedang, dan sangat resistan, masing-masing, dalam satu populasi GR waterhemp dari Kansas. Chatham dkk. ( 2015 ) melaporkan peningkatan jumlah salinan gen EPSPS (2 hingga 12 kali lipat) pada populasi GR waterhemp dari Kansas, Nebraska, Illinois, dan Missouri. Dalam studi yang sama, beberapa tanaman waterhemp dari keempat negara bagian yang bertahan hidup pada tingkat glifosat yang lebih tinggi (1680 atau 3360 g ae ha −1 ) tidak mengalami amplifikasi gen EPSPS dan kemungkinan mekanisme resistensi alternatif diajukan (Chatham dkk., 2015 ).
4 KESIMPULAN DAN IMPLIKASI PRAKTIS
Hasil ini melaporkan kasus pertama GR waterhemp di Connecticut. Biotipe CT_Res waterhemp menunjukkan tingkat resistensi sedang (5,8 kali lipat) dan amplifikasi gen EPSPS diamati pada salah satu dari tiga tanaman GR yang diuji. Tidak adanya mekanisme target-situs (mutasi titik pada gen EPSPS atau amplifikasi gen EPSPS ) pada dua tanaman GR waterhemp lain yang dikonfirmasi memerlukan penyelidikan lebih lanjut terhadap mekanisme resistensi glifosat berbasis situs non-target pada biotipe GR waterhemp dari CT. Petani disarankan untuk menggunakan beberapa taktik pengendalian untuk mengelola dan mencegah penyebaran lebih lanjut dari biotipe GR waterhemp. Pengintegrasian pengolahan tanah, tanaman penutup, dan herbisida alternatif pra-tumbuh dan pasca-tumbuh dengan beberapa MOA telah dilaporkan memberikan pengendalian gulma yang resistan terhadap herbisida sepanjang musim (Aulakh et al., 2012, 2013 ; Jhala et al., 2014 ; Kumar et al., 2025 ; Norsworthy et al., 2012 ). Penelitian mendatang akan mengevaluasi taktik pengelolaan gulma terpadu (IWM), termasuk penggunaan herbisida pra-tumbuh dan pasca-tumbuh yang efektif dalam kombinasi dengan tanaman penutup, rotasi tanaman kompetitif, pengolahan tanah, dan teknik pengendalian benih gulma panen (HWSC) untuk mengelola gulma waterhemp GR dan mencegah penyebarannya di CT dan wilayah timur laut.



Tinggalkan Balasan